

The role of shear stress in Blood-Brain Barrier endothelial physiology
- PMID: 21569296
- PMCID: PMC3103473
- DOI: 10.1186/1471-2202-12-40
The role of shear stress in Blood-Brain Barrier endothelial physiology
Abstract
Background: One of the most important and often neglected physiological stimuli contributing to the differentiation of vascular endothelial cells (ECs) into a blood-brain barrier (BBB) phenotype is shear stress (SS). With the use of a well established humanized dynamic in vitro BBB model and cDNA microarrays, we have profiled the effect of SS in the induction/suppression of ECs genes and related functions.
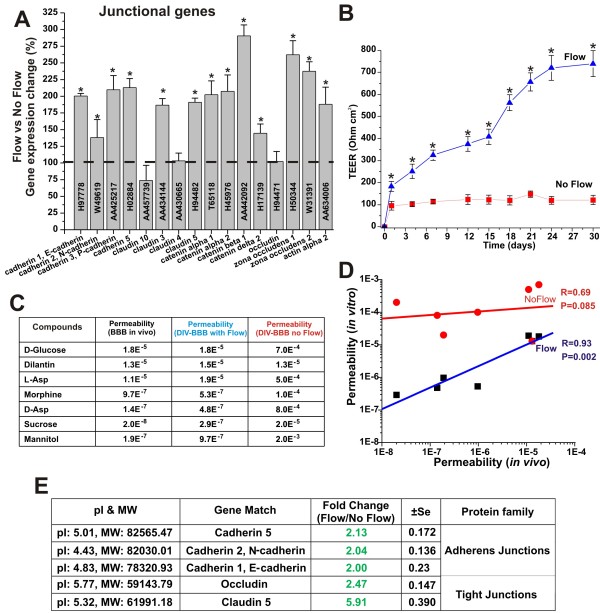
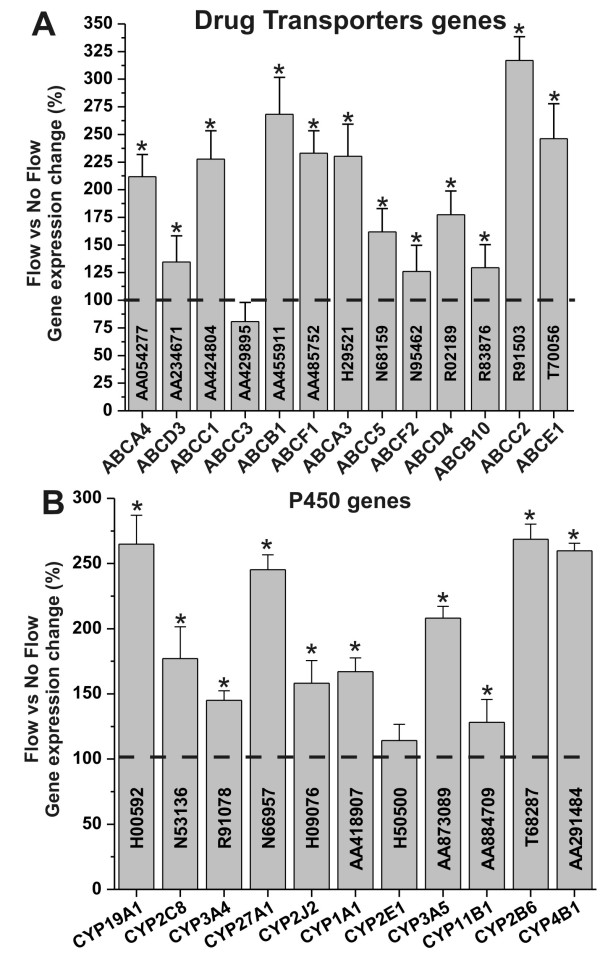
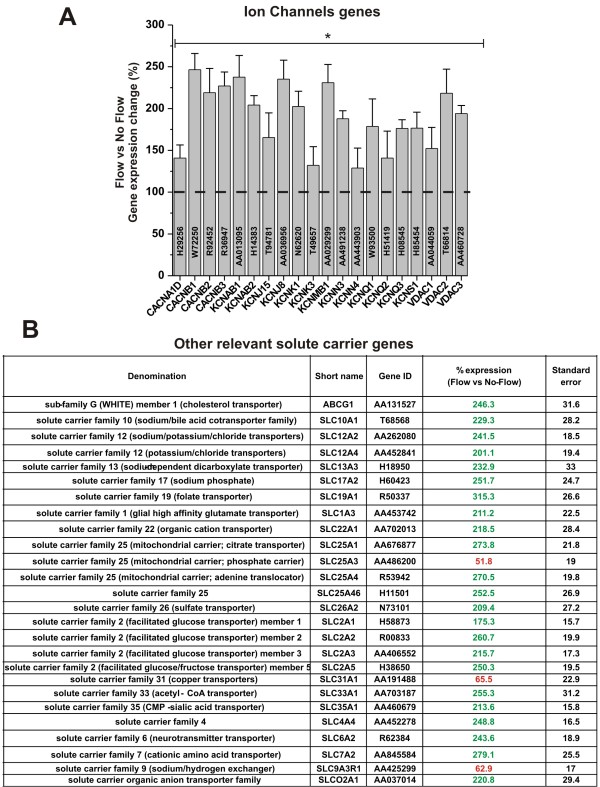
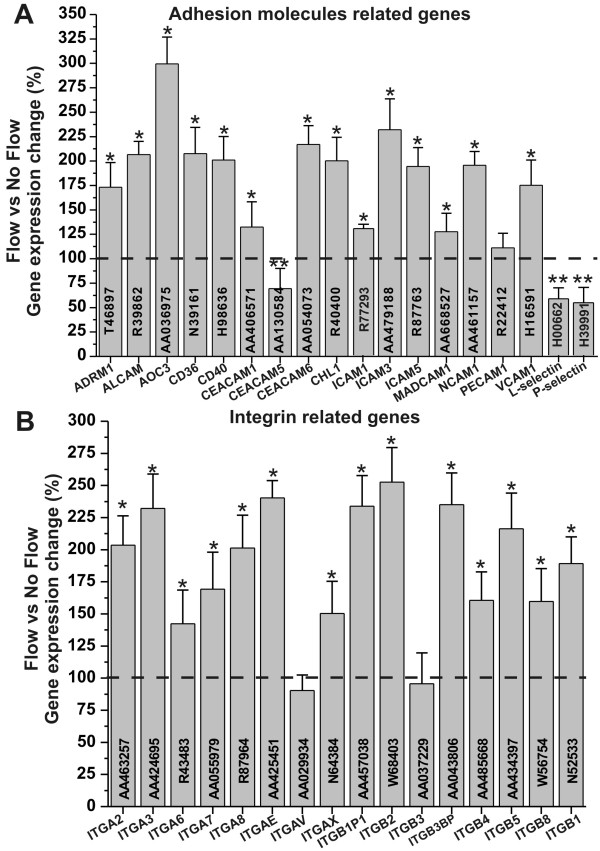
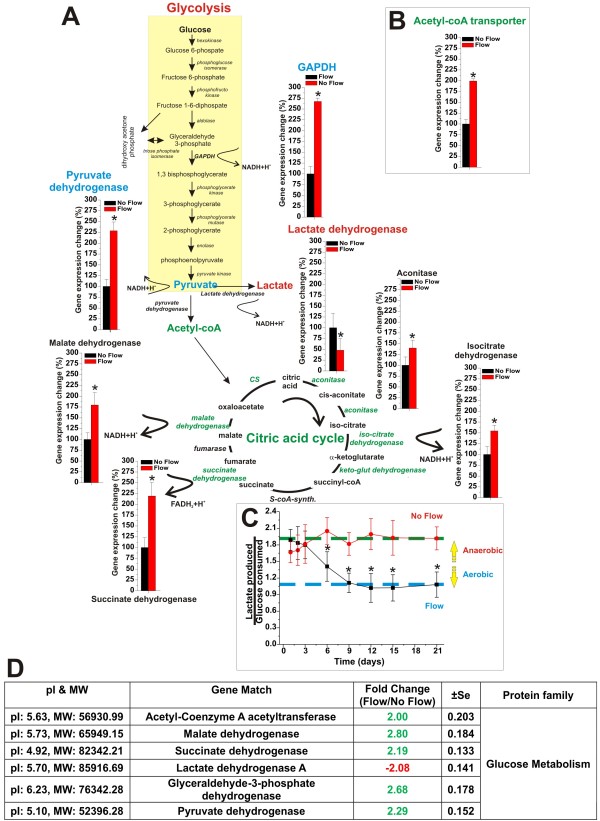
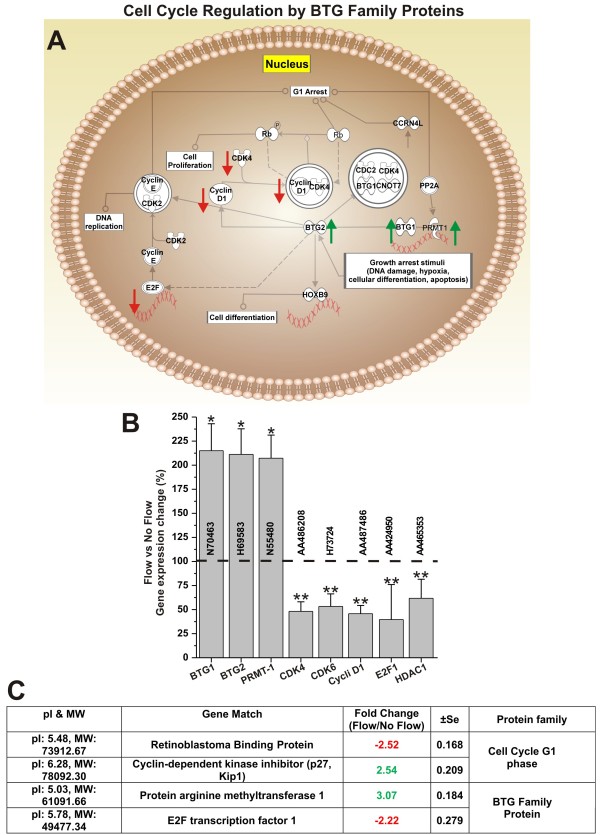
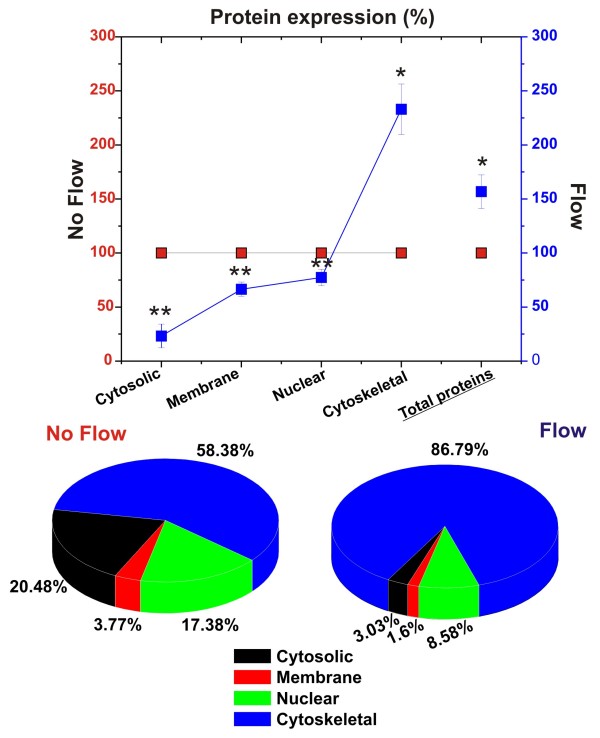
Results: Specifically, we found a significant upregulation of tight and adherens junctions proteins and genes. Trans-endothelial electrical resistance (TEER) and permeability measurements to know substances have shown that SS promoted the formation of a tight and highly selective BBB. SS also increased the RNA level of multidrug resistance transporters, ion channels, and several p450 enzymes. The RNA level of a number of specialized carrier-mediated transport systems (e.g., glucose, monocarboxylic acid, etc.) was also upregulated.RNA levels of modulatory enzymes of the glycolytic pathway (e.g., lactate dehydrogenase) were downregulated by SS while those involved in the Krebs cycle (e.g., lactate and other dehydrogenases) were upregulated. Measurements of glucose consumption versus lactate production showed that SS negatively modulated the glycolytic bioenergetic pathways of glucose metabolism in favor of the more efficient aerobic respiration. BBB ECs are responsive to inflammatory stimuli. Our data showed that SS increased the RNA levels of integrins and vascular adhesion molecules. SS also inhibited endothelial cell cycle via regulation of BTG family proteins encoding genes. This was paralleled by significant increase in the cytoskeletal protein content while that of membrane, cytosol, and nuclear sub-cellular fractions decreased. Furthermore, analysis of 2D gel electrophoresis (which allows identifying a large number of proteins per sample) of EC proteins extracted from membrane sub-cellular endothelial fractions showed that SS increased the expression levels of tight junction proteins. In addition, regulatory enzymes of the Krebb's cycle (aerobic glucose metabolism) were also upregulated. Furthermore, the expression pattern of key protein regulators of the cell cycle and parallel gene array data supported a cell proliferation inhibitory role for SS.
Conclusions: Genomic and proteomic analyses are currently used to examine BBB function in healthy and diseased brain and characterize this dynamic interface. In this study we showed that SS plays a key role in promoting the differentiation of vascular endothelial cells into a truly BBB phenotype. SS affected multiple aspect of the endothelial physiology spanning from tight junctions formation to cell division as well as the expression of multidrug resistance transporters. BBB dysfunction has been observed in many neurological diseases, but the causes are generally unknown. Our study provides essential insights to understand the role played by SS in the BBB formation and maintenance.
Figures
References
-
- Steiner O, Coisne C, Cecchelli R, Boscacci R, Deutsch U, Engelhardt B. et al. Differential roles for endothelial ICAM-1, ICAM-2, and VCAM-1 in shear-resistant T cell arrest, polarization, and directed crawling on blood-brain barrier endothelium. J Immunol. 2010;185:4846–4855. doi: 10.4049/jimmunol.0903732. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
