

Reversible response of protein localization and microtubule organization to nutrient stress during Drosophila early oogenesis
- PMID: 21570389
- PMCID: PMC3118931
- DOI: 10.1016/j.ydbio.2011.04.022
Reversible response of protein localization and microtubule organization to nutrient stress during Drosophila early oogenesis
Abstract
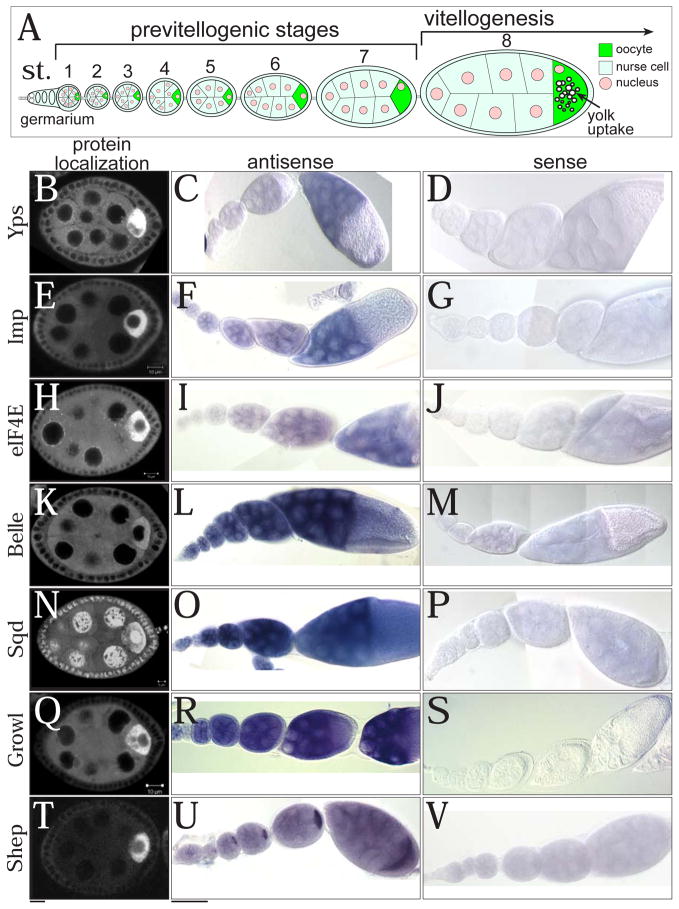
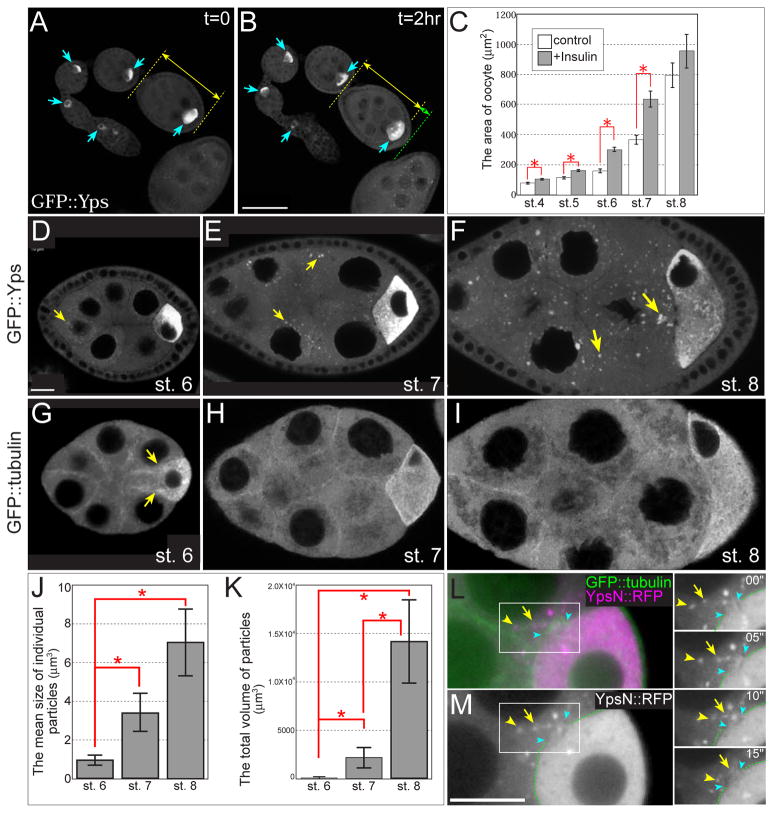
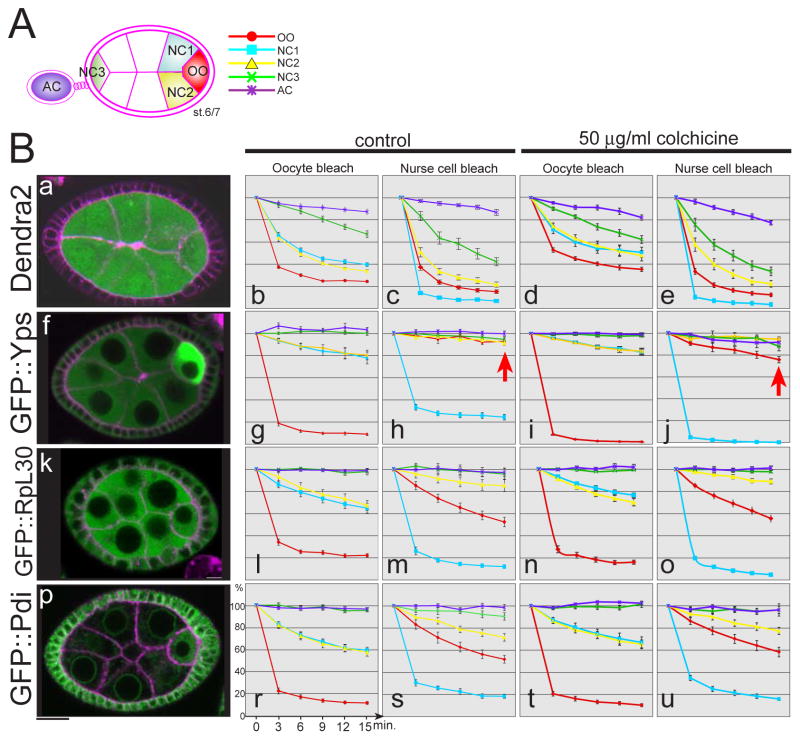
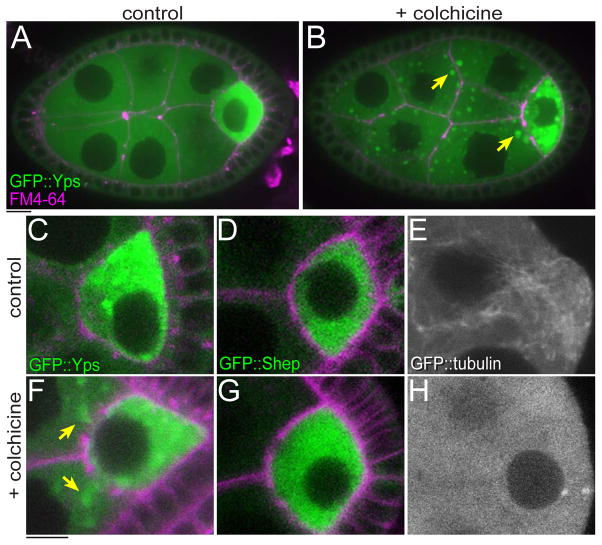
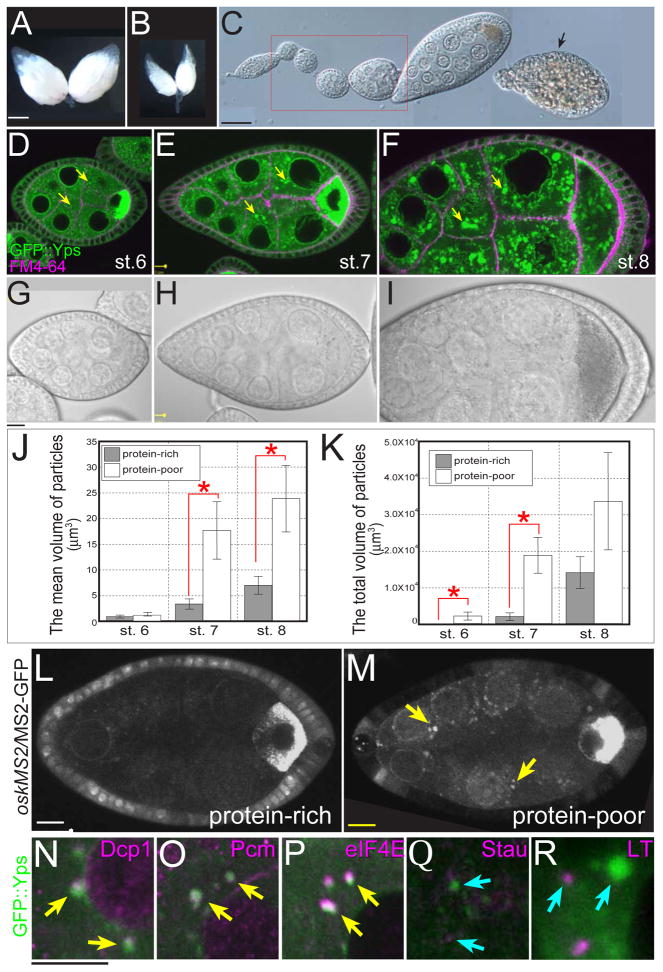
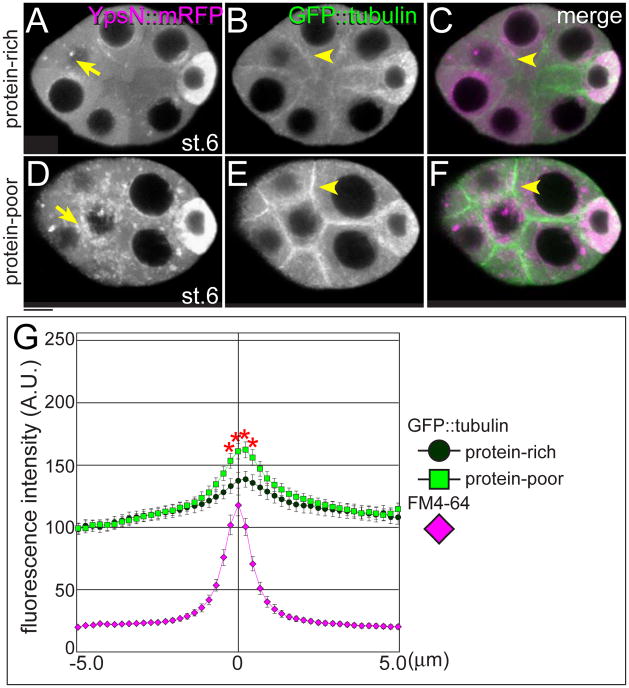
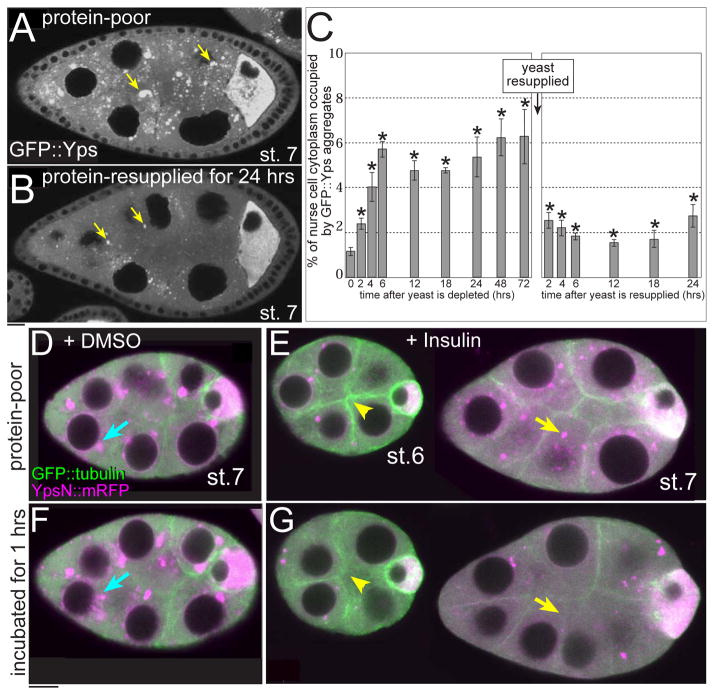
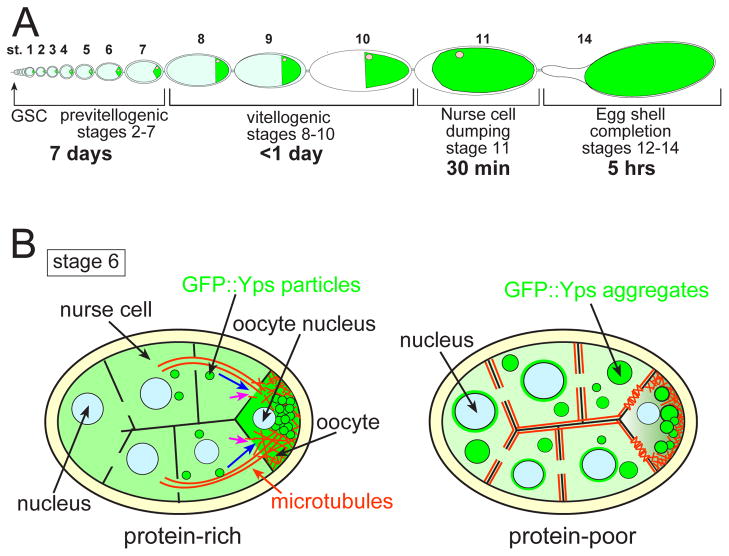
The maturation of animal oocytes is highly sensitive to nutrient availability. During Drosophila oogenesis, a prominent metabolic checkpoint occurs at the onset of yolk uptake (vitellogenesis): under nutrient stress, egg chambers degenerate by apoptosis. To investigate additional responses to nutrient deprivation, we studied the intercellular transport of cytoplasmic components between nurse cells and the oocyte during previtellogenic stages. Using GFP protein-traps, we showed that Ypsilon Schachtel (Yps), a putative RNA binding protein, moved into the oocyte by both microtubule (MT)-dependent and -independent mechanisms, and was retained in the oocyte in a MT-dependent manner. These data suggest that oocyte enrichment is accomplished by a combination of MT-dependent polarized transport and MT-independent flow coupled with MT-dependent trapping within the oocyte. Under nutrient stress, Yps and other components of the oskar ribonucleoprotein complex accumulated in large processing bodies in nurse cells, accompanied by MT reorganization. This response was detected as early as 2h after starvation, suggesting that young egg chambers rapidly respond to nutrient stress. Moreover, both Yps aggregation and MT reorganization were reversed with re-feeding of females or the addition of exogenous insulin to cultured egg chambers. Our results suggest that egg chambers rapidly mount a stress response by altering intercellular transport upon starvation. This response implies a mechanism for preserving young egg chambers so that egg production can rapidly resume when nutrient availability improves.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Anderson P, Kedersha N. Stress granules: the Tao of RNA triage. Trends Biochem Sci. 2008;33:141–50. - PubMed
-
- Anderson P, Kedersha N. RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol. 2009;10:430–6. - PubMed
-
- Ashburner M. Drosophila. Cold Spring Harbor Laboratory; Cold Spring Harbor, N.Y: 1989.
-
- Bohrmann J, Biber K. Cytoskeleton-dependent transport of cytoplasmic particles in previtellogenic to mid-vitellogenic ovarian follicles of Drosophila: time-lapse analysis using video-enhanced contrast microscopy. J Cell Sci. 1994;107(Pt 4):849–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
