

Genetic analysis of age-dependent defects of the Caenorhabditis elegans touch receptor neurons
- PMID: 21571636
- PMCID: PMC3107274
- DOI: 10.1073/pnas.1011711108
Genetic analysis of age-dependent defects of the Caenorhabditis elegans touch receptor neurons
Abstract
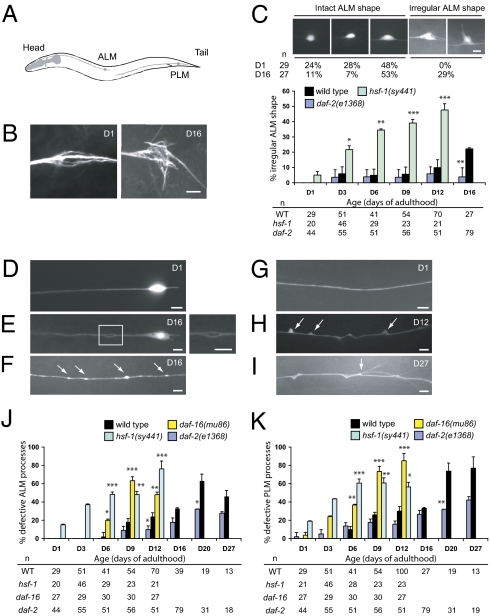
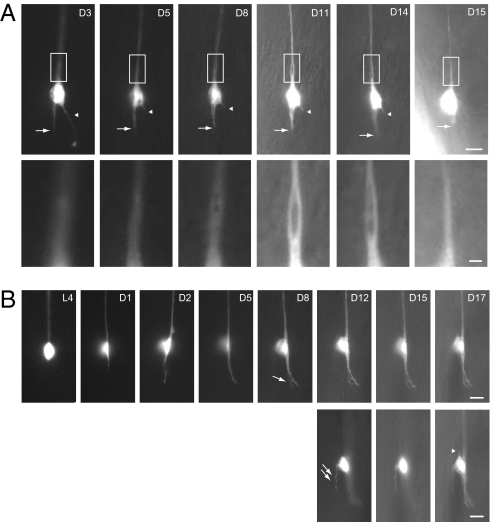
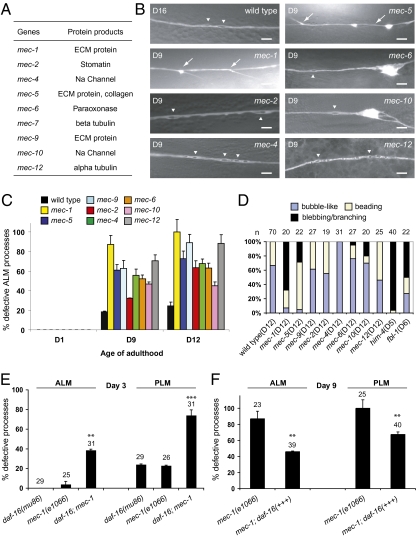
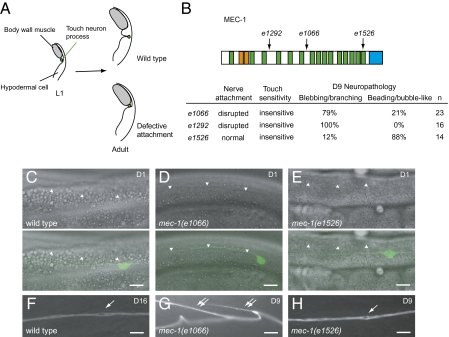
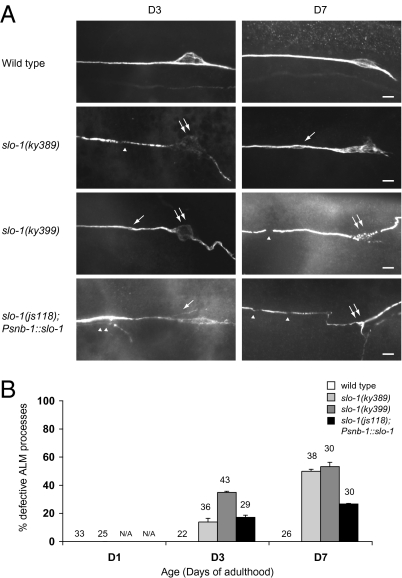
Although many genes have been implicated in the pathogenesis of common neurodegenerative diseases, the genetic and cellular mechanisms that maintain neuronal integrity during normal aging remain elusive. Here we show that Caenorhabditis elegans touch receptor and cholinergic neurons display age-dependent morphological defects, including cytoskeletal disorganization, axon beading, and defasciculation. Progression of neuronal aging is regulated by DAF-2 and DAF-16 signaling, which also modulate adult life span. Mutations that disrupt touch-evoked sensory activity or reduce membrane excitability trigger accelerated neuronal aging, indicating that electrical activity is critical for adult neuronal integrity. Disrupting touch neuron attachment to the epithelial cells induces distinct neurodegenerative phenotypes. These results provide a detailed description of the age-dependent morphological defects that occur in identified neurons of C. elegans, demonstrate that the age of onset of these defects is regulated by specific genes, and offer experimental evidence for the importance of normal levels of neural activity in delaying neuronal aging.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
