

Temporal encoding in a nervous system
- PMID: 21573206
- PMCID: PMC3088658
- DOI: 10.1371/journal.pcbi.1002041
Temporal encoding in a nervous system
Abstract
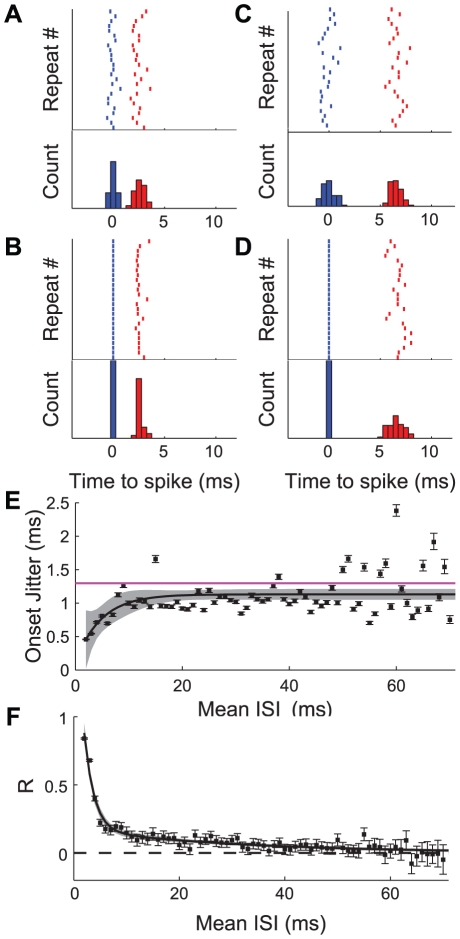
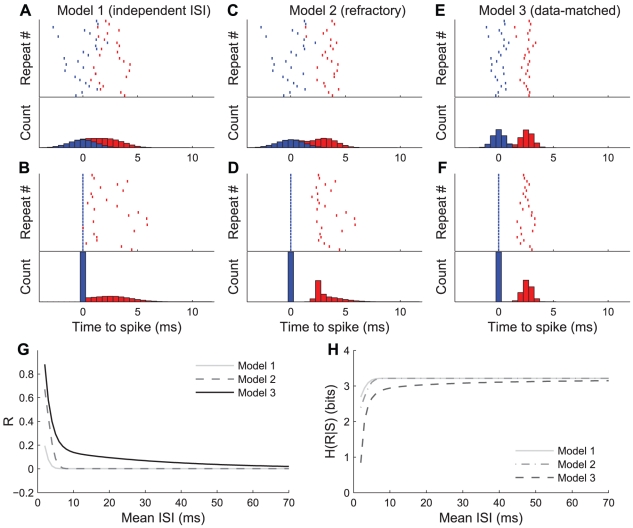
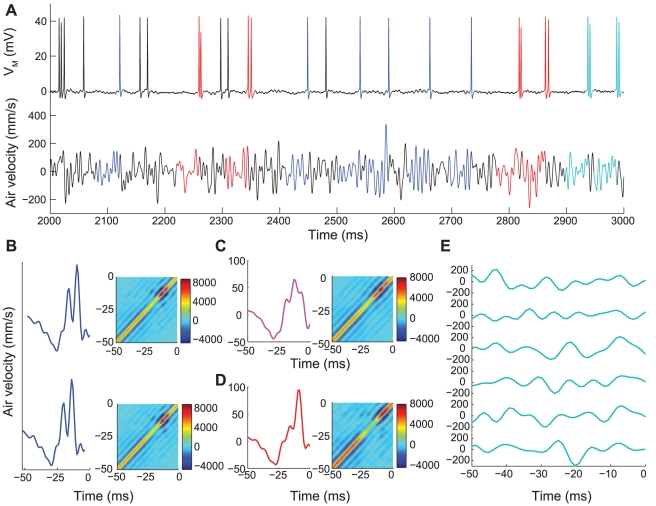
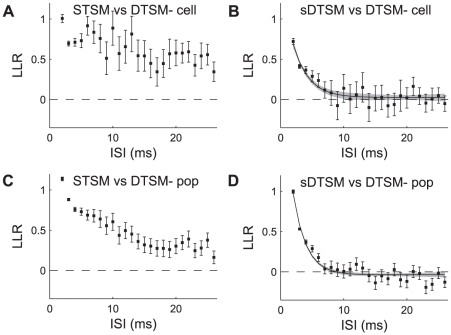
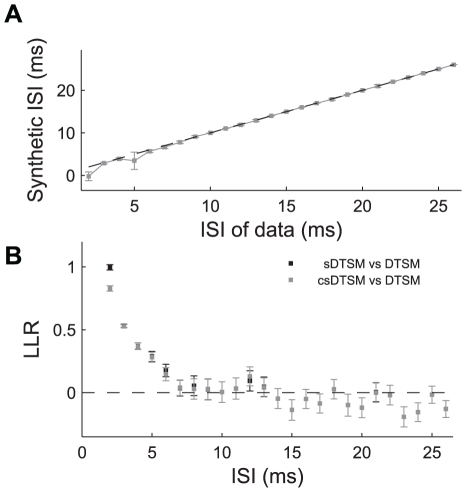
We examined the extent to which temporal encoding may be implemented by single neurons in the cercal sensory system of the house cricket Acheta domesticus. We found that these neurons exhibit a greater-than-expected coding capacity, due in part to an increased precision in brief patterns of action potentials. We developed linear and non-linear models for decoding the activity of these neurons. We found that the stimuli associated with short-interval patterns of spikes (ISIs of 8 ms or less) could be predicted better by second-order models as compared to linear models. Finally, we characterized the difference between these linear and second-order models in a low-dimensional subspace, and showed that modification of the linear models along only a few dimensions improved their predictive power to parity with the second order models. Together these results show that single neurons are capable of using temporal patterns of spikes as fundamental symbols in their neural code, and that they communicate specific stimulus distributions to subsequent neural structures.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Information transmission in cercal giant interneurons is unaffected by axonal conduction noise.PLoS One. 2012;7(1):e30115. doi: 10.1371/journal.pone.0030115. Epub 2012 Jan 12. PLoS One. 2012. PMID: 22253900 Free PMC article.
-
Information theoretic analysis of dynamical encoding by four identified primary sensory interneurons in the cricket cercal system.J Neurophysiol. 1996 Apr;75(4):1345-64. doi: 10.1152/jn.1996.75.4.1345. J Neurophysiol. 1996. PMID: 8727382
-
Encoding and decoding spikes for dynamic stimuli.Neural Comput. 2008 Sep;20(9):2325-60. doi: 10.1162/neco.2008.01-07-436. Neural Comput. 2008. PMID: 18386986
-
Computational mechanisms of mechanosensory processing in the cricket.J Exp Biol. 2008 Jun;211(Pt 11):1819-28. doi: 10.1242/jeb.016402. J Exp Biol. 2008. PMID: 18490398 Review.
-
The neuronal encoding of information in the brain.Prog Neurobiol. 2011 Nov;95(3):448-90. doi: 10.1016/j.pneurobio.2011.08.002. Epub 2011 Sep 2. Prog Neurobiol. 2011. PMID: 21907758 Review.
Cited by
-
Action selection based on multiple-stimulus aspects in wind-elicited escape behavior of crickets.Heliyon. 2022 Jan 20;8(1):e08800. doi: 10.1016/j.heliyon.2022.e08800. eCollection 2022 Jan. Heliyon. 2022. PMID: 35111985 Free PMC article.
-
Encoding of small-scale air motion dynamics in the cricket, Acheta domesticus.J Neurophysiol. 2022 Apr 1;127(4):1185-1197. doi: 10.1152/jn.00042.2022. Epub 2022 Mar 30. J Neurophysiol. 2022. PMID: 35353628 Free PMC article.
-
A minimum-error, energy-constrained neural code is an instantaneous-rate code.J Comput Neurosci. 2016 Apr;40(2):193-206. doi: 10.1007/s10827-016-0592-x. Epub 2016 Feb 27. J Comput Neurosci. 2016. PMID: 26922680
-
Symmetry-Breaking Bifurcations of the Information Bottleneck and Related Problems.Entropy (Basel). 2022 Sep 2;24(9):1231. doi: 10.3390/e24091231. Entropy (Basel). 2022. PMID: 36141117 Free PMC article.
-
Capturing spike train temporal pattern with wavelet average coefficient for brain machine interface.Sci Rep. 2021 Sep 24;11(1):19020. doi: 10.1038/s41598-021-98578-5. Sci Rep. 2021. PMID: 34561503 Free PMC article.
References
-
- Bialek W, Rieke F, de Ruyter van Steveninck RR, Warland D. Reading a neural code. Science. 1991;252:1854–1857. - PubMed
-
- Rieke F, Warland D, Bialek W, de Ruyter van Steveninck RR. Spikes: exploring the neural code. Cambridge, Mass. London: MIT Press; 1997. 416
-
- Theunissen F, Miller JP. Temporal encoding in nervous systems: a rigorous definition. J Comput Neurosci. 1995;2:149–162. - PubMed
-
- Theunissen F, Roddey JC, Stufflebeam S, Clague H, Miller JP. Information theoretic analysis of dynamical encoding by four identified primary sensory interneurons in the cricket cercal system. J Neurophysiol. 1996;75:1345–1364. - PubMed
-
- de Ruyter van Steveninck RR, Lewen GD, Strong SP, Koberle R, Bialek W. Reproducibility and variability in neural spike trains. Science. 1997;275:1805–1808. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
