

The interaction between two extracellular linker regions controls sustained opening of acid-sensing ion channel 1
- PMID: 21576243
- PMCID: PMC3129216
- DOI: 10.1074/jbc.M111.230797
The interaction between two extracellular linker regions controls sustained opening of acid-sensing ion channel 1
Abstract
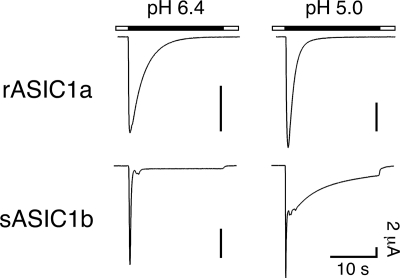
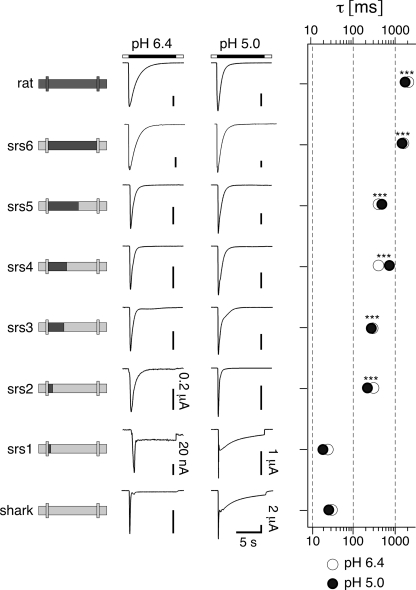
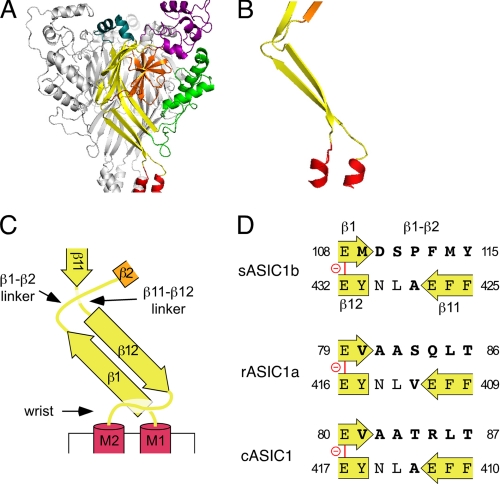
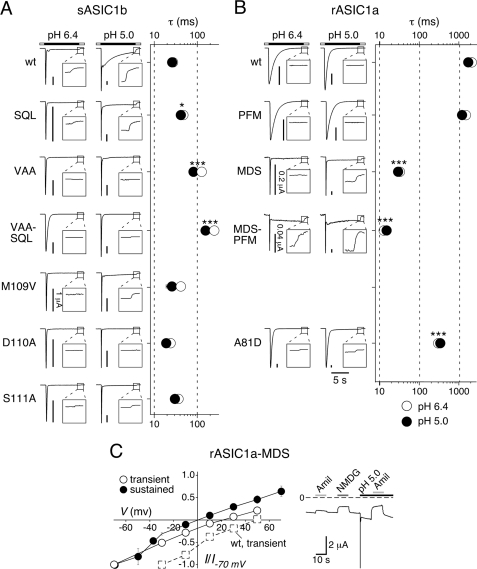
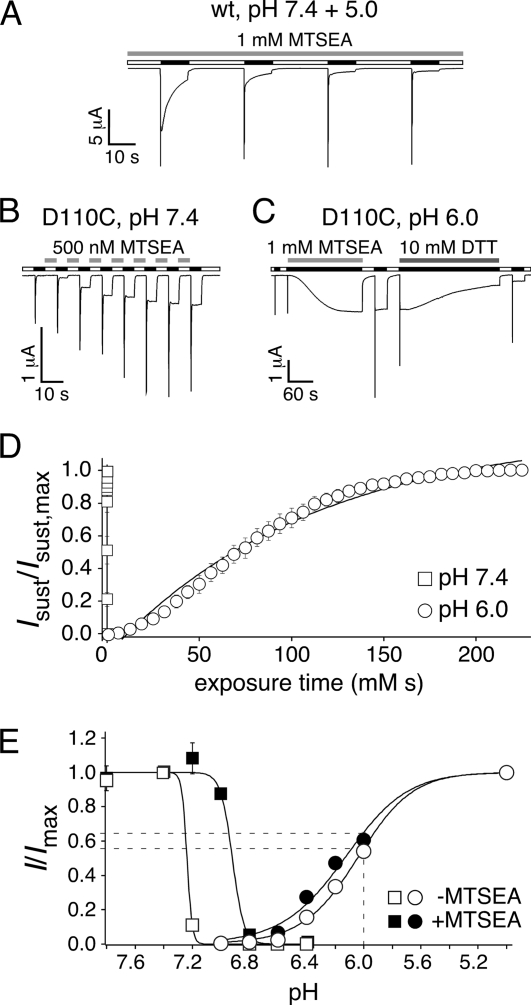
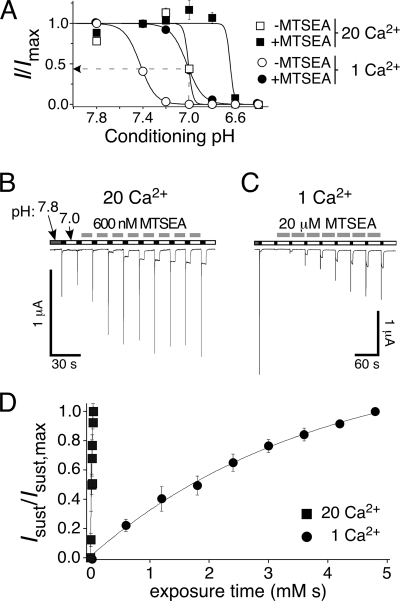
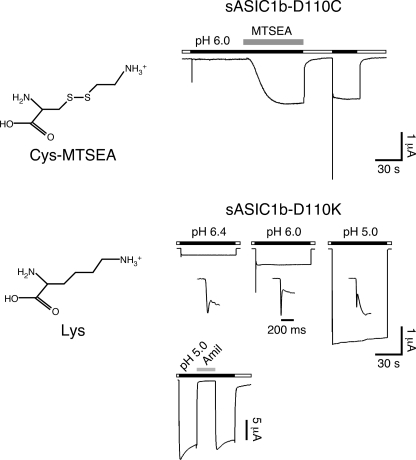
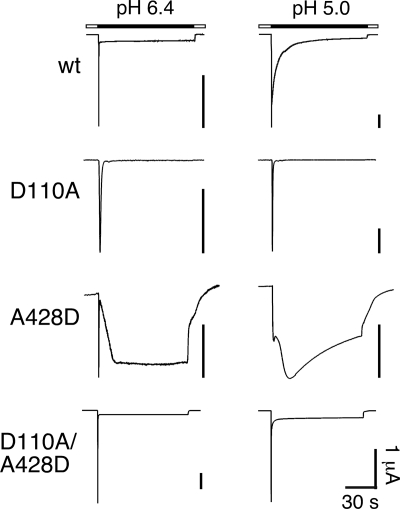
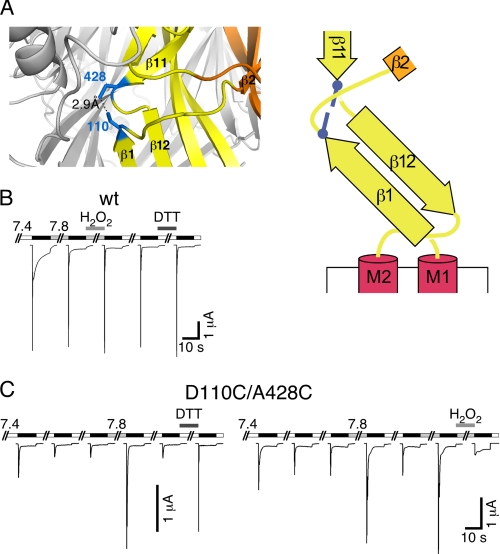
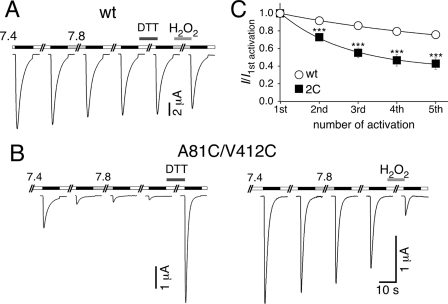
Activation of acid-sensing ion channels (ASICs) contributes to neuronal death during stroke, to axonal degeneration during neuroinflammation, and to pain during inflammation. Although understanding ASIC gating may help to modulate ASIC activity during these pathologic situations, at present it is poorly understood. The ligand, H(+), probably binds to several sites, among them amino acids within the large extracellular domain. The extracellular domain is linked to the two transmembrane domains by the wrist region that is connected to two anti-parallel β-strands, β1 and β12. Thus, the wrist region together with those β-strands may have a crucial role in transmitting ligand binding to pore opening and closing. Here we show that amino acids in the β1-β2 linker determine constitutive opening of ASIC1b from shark. The most crucial residue within the β1-β2 linker (Asp(110)), when mutated from aspartate to cysteine, can be altered by cysteine-modifying reagents much more readily when channels are closed than when they are desensitized. Finally, engineering of a cysteine at position 110 and at an adjacent position in the β11-β12 linker leads to spontaneous formation of a disulfide bond that traps the channel in the desensitized conformation. Collectively, our results suggest that the β1-β2 and β11-β12 linkers are dynamic during gating and tightly appose to each other during desensitization gating. Hindrance of this tight apposition leads to reopening of the channel. It follows that the β1-β2 and β11-β12 linkers modulate gating movements of ASIC1 and may thus be drug targets to modulate ASIC activity.
Figures
References
-
- Wemmie J. A., Price M. P., Welsh M. J. (2006) Trends Neurosci. 29, 578–586 - PubMed
-
- Askwith C. C., Wemmie J. A., Price M. P., Rokhlina T., Welsh M. J. (2004) J. Biol. Chem. 279, 18296–18305 - PubMed
-
- Xiong Z. G., Zhu X. M., Chu X. P., Minami M., Hey J., Wei W. L., MacDonald J. F., Wemmie J. A., Price M. P., Welsh M. J., Simon R. P. (2004) Cell 118, 687–698 - PubMed
-
- Wemmie J. A., Chen J., Askwith C. C., Hruska-Hageman A. M., Price M. P., Nolan B. C., Yoder P. G., Lamani E., Hoshi T., Freeman J. H., Jr., Welsh M. J. (2002) Neuron 34, 463–477 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
