

Phosphoinositides in cell architecture
- PMID: 21576256
- PMCID: PMC3140688
- DOI: 10.1101/cshperspect.a004796
Phosphoinositides in cell architecture
Abstract
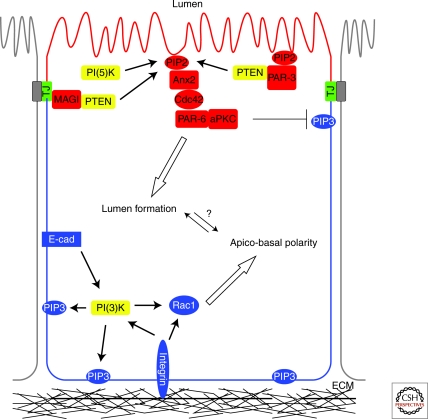
Inositol phospholipids have been implicated in almost all aspects of cellular physiology including spatiotemporal regulation of cellular signaling, acquisition of cellular polarity, specification of membrane identity, cytoskeletal dynamics, and regulation of cellular adhesion, motility, and cytokinesis. In this review, we examine the critical role phosphoinositides play in these processes to execute the establishment and maintenance of cellular architecture. Epithelial tissues perform essential barrier and transport functions in almost all major organs. Key to their development and function is the establishment of epithelial cell polarity. We place a special emphasis on highlighting recent studies demonstrating phosphoinositide regulation of epithelial cell polarity and how individual cells use phosphoinositides to further organize into epithelial tissues.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources