

Limiting Ago protein restricts RNAi and microRNA biogenesis during early development in Xenopus laevis
- PMID: 21576259
- PMCID: PMC3110951
- DOI: 10.1101/gad.2038811
Limiting Ago protein restricts RNAi and microRNA biogenesis during early development in Xenopus laevis
Abstract
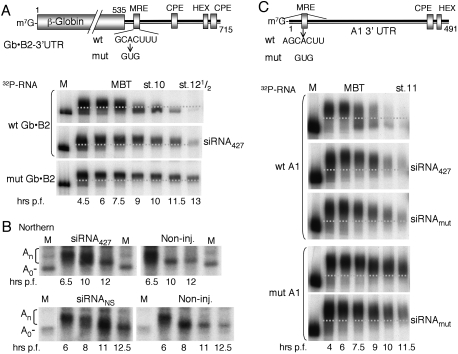
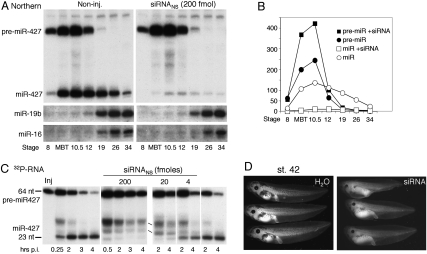
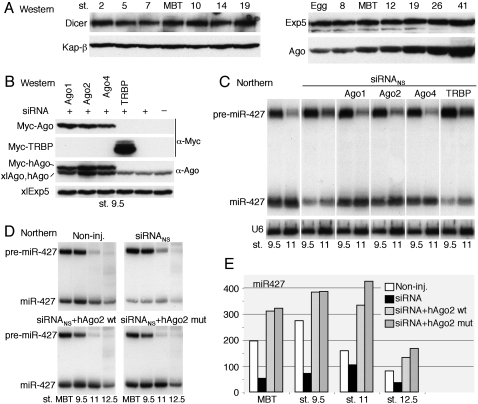
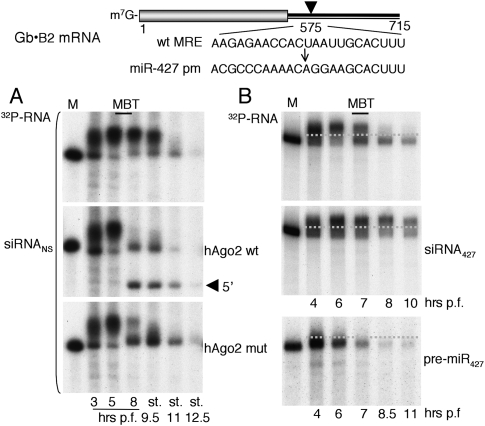
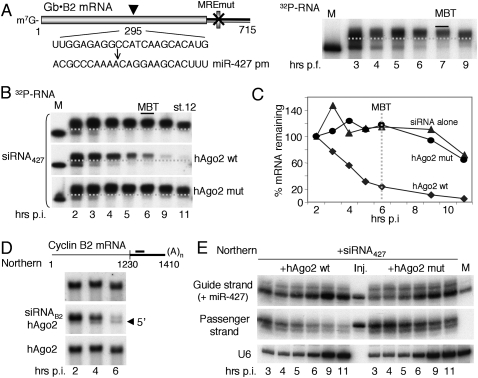
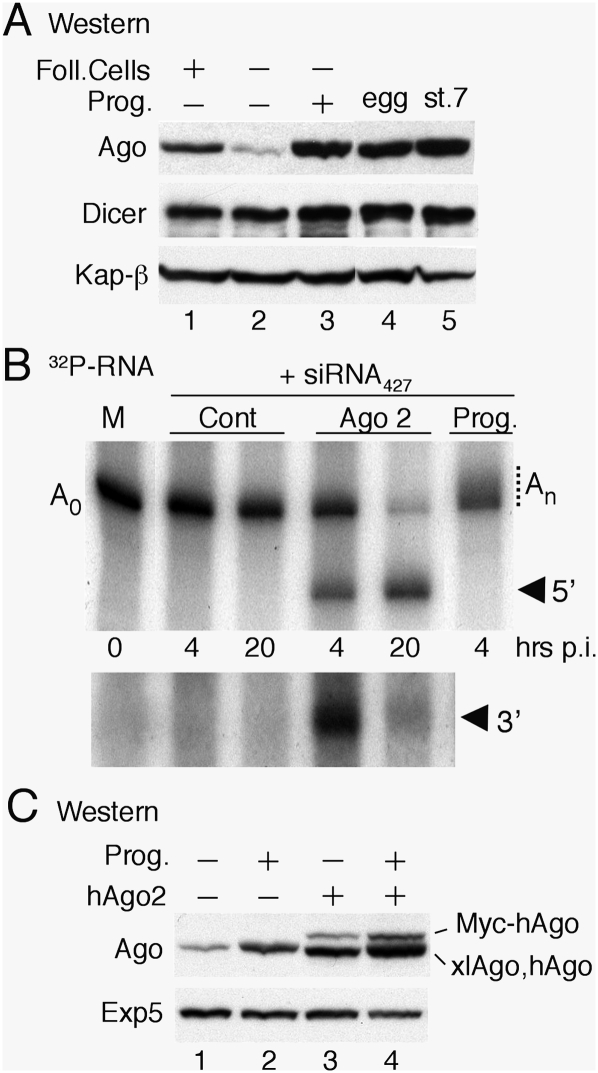
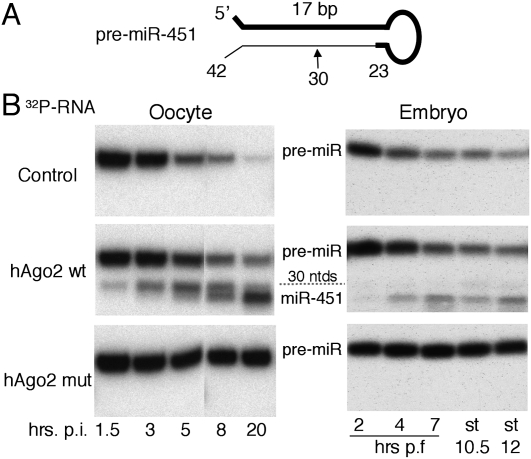
We show that, in Xenopus laevis oocytes and early embryos, double-stranded exogenous siRNAs cannot function as microRNA (miRNA) mimics in either deadenylation or guided mRNA cleavage (RNAi). Instead, siRNAs saturate and inactivate maternal Argonaute (Ago) proteins, which are present in low amounts but are needed for Dicer processing of pre-miRNAs at the midblastula transition (MBT). Consequently, siRNAs impair accumulation of newly made miRNAs, such as the abundant embryonic pre-miR-427, but inhibition dissipates upon synthesis of zygotic Ago proteins after MBT. These effects of siRNAs, which are independent of sequence, result in morphological defects at later stages of development. The expression of any of several exogenous human Ago proteins, including catalytically inactive Ago2 (Ago2mut), can overcome the siRNA-mediated inhibition of miR-427 biogenesis and function. However, expression of wild-type, catalytically active hAgo2 is required to elicit RNAi in both early embryos and oocytes using either siRNA or endogenous miRNAs as guides. The lack of endogenous Ago2 endonuclease activity explains why these cells normally are unable to support RNAi. Expression of catalytically active exogenous Ago2, which appears not to perturb normal Xenopus embryonic development, can now be exploited for RNAi in this vertebrate model organism.
Figures
Comment in
-
RNAi in Xenopus: look before you leap.Genes Dev. 2011 Jun 1;25(11):1105-8. doi: 10.1101/gad.2062911. Genes Dev. 2011. PMID: 21632820 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources