

Circadian oscillation of gibberellin signaling in Arabidopsis
- PMID: 21576475
- PMCID: PMC3107313
- DOI: 10.1073/pnas.1101050108
Circadian oscillation of gibberellin signaling in Arabidopsis
Abstract
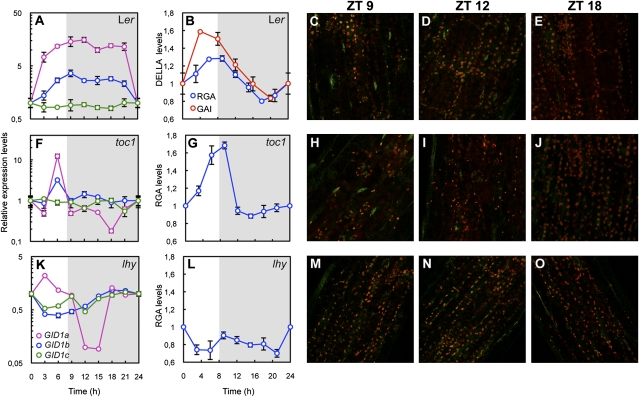
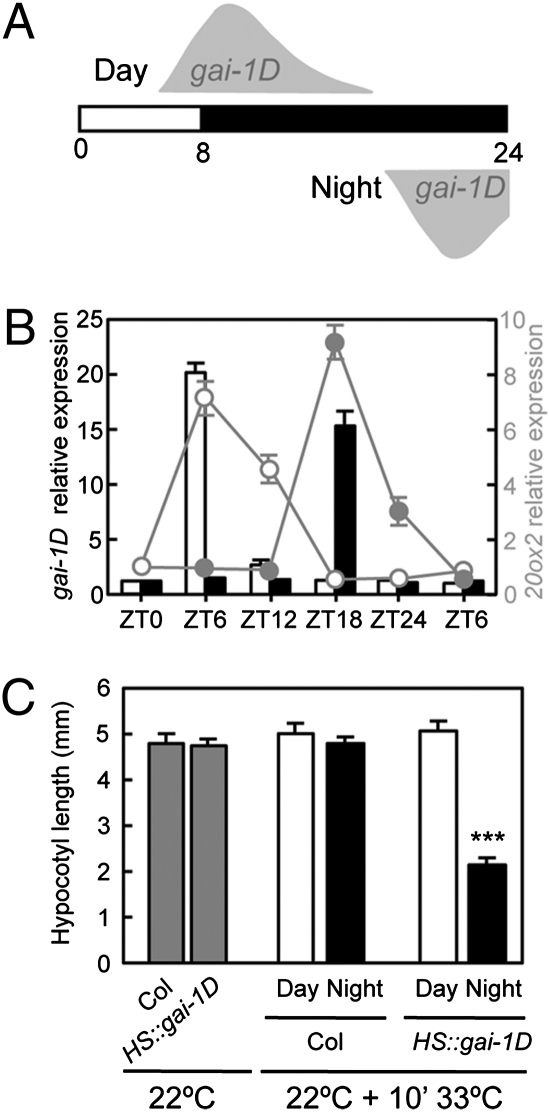
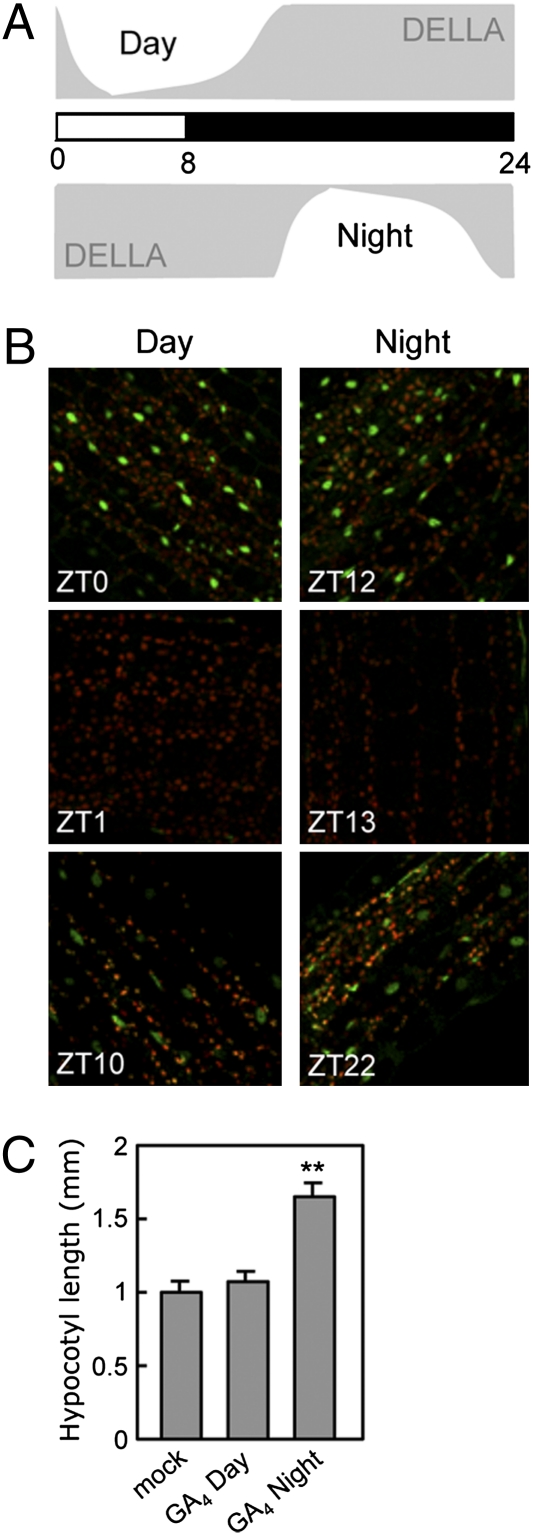
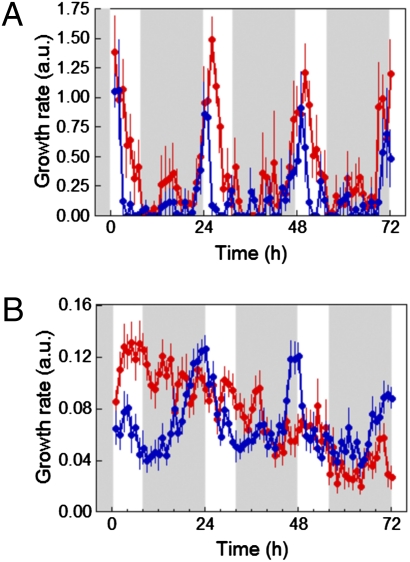
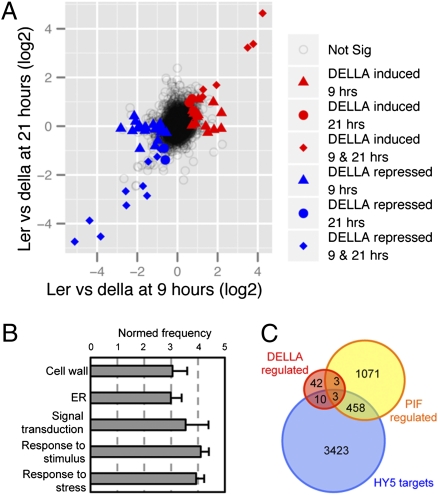
Circadian clocks are endogenous timekeeping mechanisms that allow organisms to anticipate rhythmic, daily environmental changes. Temporal coordination of transcription results in a set of gene expression patterns with peak levels occurring at precise times of the day. An intriguing question is how a single clock can generate different oscillatory rhythms, and it has been proposed that hormone signaling might act in plants as a relay mechanism to modulate the amplitude and the phase of output rhythms. Here we show that the circadian clock gates gibberellin (GA) signaling through transcriptional regulation of the GA receptors, resulting in higher stability of DELLA proteins during daytime and higher GA sensitivity at night. Oscillation of GA signaling appears to be particularly critical for rhythmic growth, given that constitutive expression of the GA receptor expands the daily growth period in seedlings, and complete loss of DELLA function causes continuous, arrhythmic hypocotyl growth. Moreover, transcriptomic analysis of a pentuple della KO mutant indicates that the GA pathway mediates the rhythmic expression of many clock-regulated genes related to biotic and abiotic stress responses and cell wall modification. Thus, gating of GA sensitivity by the circadian clock represents an additional layer of regulation that might provide extra robustness to the diurnal growth rhythm and constitute a regulatory module that coordinates the circadian clock with additional endogenous and environmental signals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nozue K, et al. Rhythmic growth explained by coincidence between internal and external cues. Nature. 2007;448:358–361. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
