

Structural topology of phospholamban pentamer in lipid bilayers by a hybrid solution and solid-state NMR method
- PMID: 21576492
- PMCID: PMC3107283
- DOI: 10.1073/pnas.1016535108
Structural topology of phospholamban pentamer in lipid bilayers by a hybrid solution and solid-state NMR method
Abstract
Phospholamban (PLN) is a type II membrane protein that inhibits the sarcoplasmic reticulum Ca(2+)-ATPase (SERCA), thereby regulating calcium homeostasis in cardiac muscle. In membranes, PLN forms pentamers that have been proposed to function either as a storage for active monomers or as ion channels. Here, we report the T-state structure of pentameric PLN solved by a hybrid solution and solid-state NMR method. In lipid bilayers, PLN adopts a pinwheel topology with a narrow hydrophobic pore, which excludes ion transport. In the T state, the cytoplasmic amphipathic helices (domains Ia) are absorbed into the lipid bilayer with the transmembrane domains arranged in a left-handed coiled-coil configuration, crossing the bilayer with a tilt angle of approximately 11° with respect to the membrane normal. The tilt angle difference between the monomer and pentamer is approximately 13°, showing that intramembrane helix-helix association forces dominate over the hydrophobic mismatch, driving the overall topology of the transmembrane assembly. Our data reveal that both topology and function of PLN are shaped by the interactions with lipids, which fine-tune the regulation of SERCA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
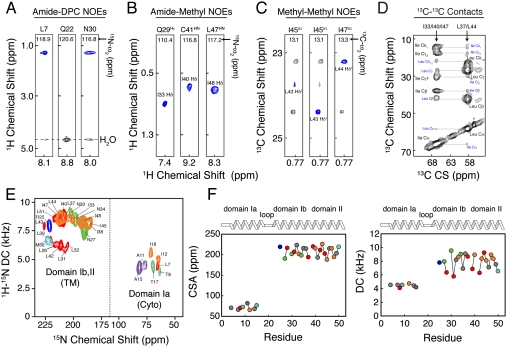
 PLN. (C) Two-dimensional planes from 3D [1H, 13C, 13C]-HSQC-NOESY-HSQC experiment performed on a sample containing 1/1 of
PLN. (C) Two-dimensional planes from 3D [1H, 13C, 13C]-HSQC-NOESY-HSQC experiment performed on a sample containing 1/1 of  and
and  PLN. (D) Two-dimensional-DARR experiment (200-ms mixing time) on a 1/1 sample of U-13C-Leu/U-13C-Ile PLN. Intraresidue and interprotomer cross-peaks are labeled in black and blue, respectively. (E) Overlay of 2D [1H, 15N]-PISEMA spectra of selectively labeled PLN in oriented DOPC/DOPE (4/1) lipid bilayers. (F) Plot of the CSA (Left) and DC (Right) values versus residue.
PLN. (D) Two-dimensional-DARR experiment (200-ms mixing time) on a 1/1 sample of U-13C-Leu/U-13C-Ile PLN. Intraresidue and interprotomer cross-peaks are labeled in black and blue, respectively. (E) Overlay of 2D [1H, 15N]-PISEMA spectra of selectively labeled PLN in oriented DOPC/DOPE (4/1) lipid bilayers. (F) Plot of the CSA (Left) and DC (Right) values versus residue.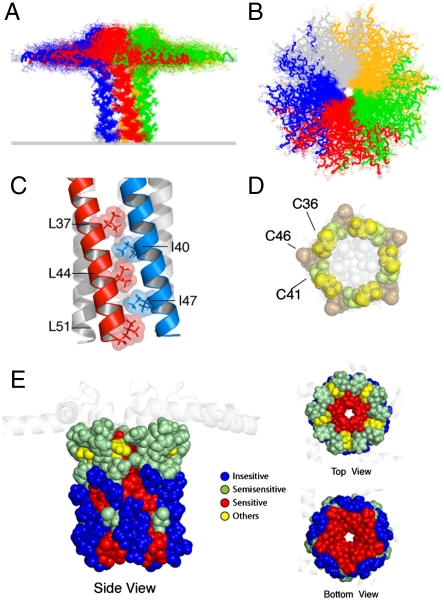
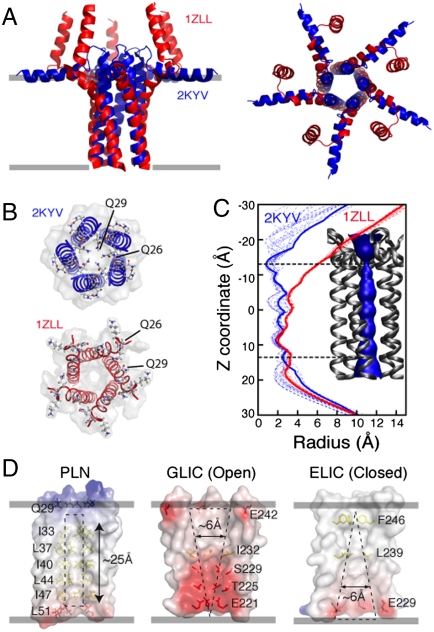
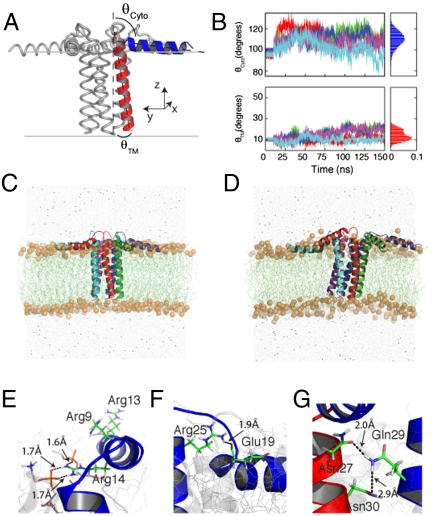
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
