

G4 resolvase 1 tightly binds and unwinds unimolecular G4-DNA
- PMID: 21586581
- PMCID: PMC3167620
- DOI: 10.1093/nar/gkr234
G4 resolvase 1 tightly binds and unwinds unimolecular G4-DNA
Abstract
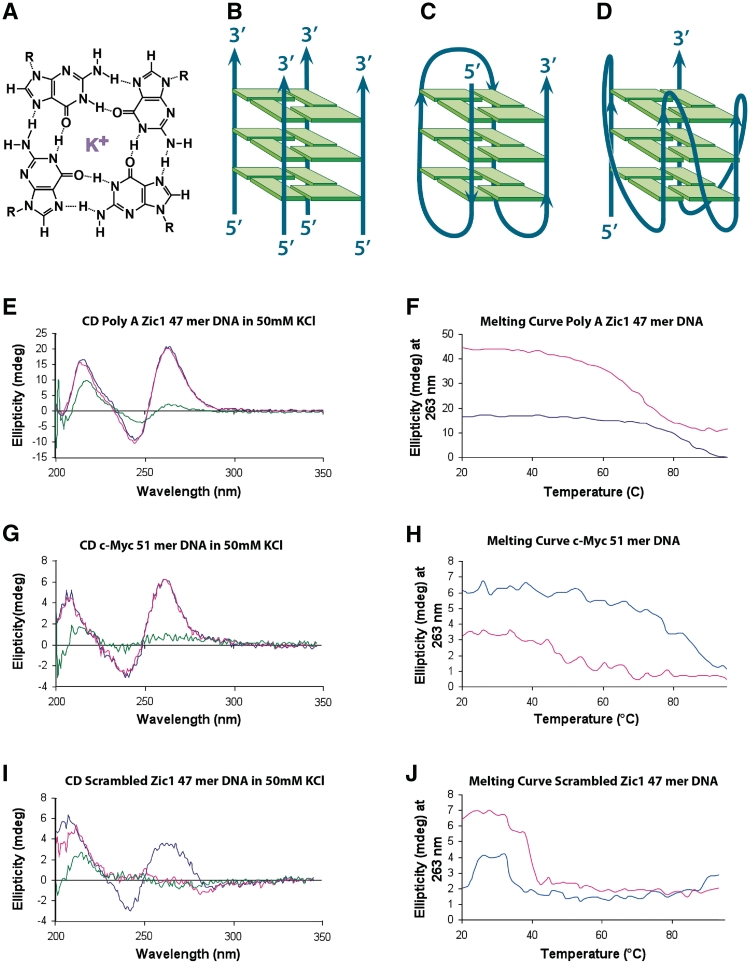
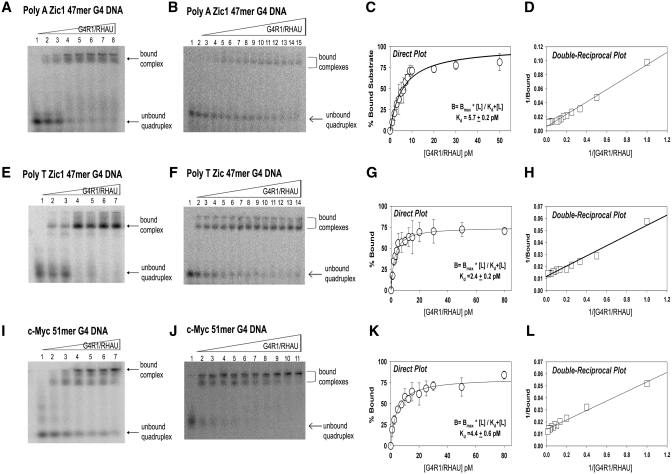
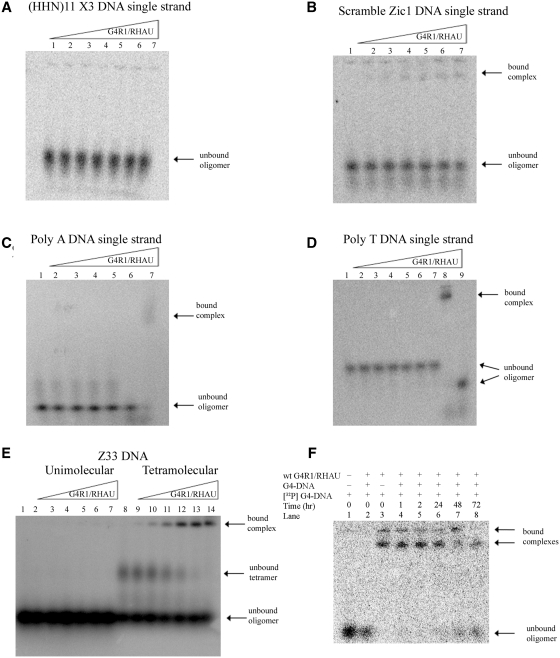
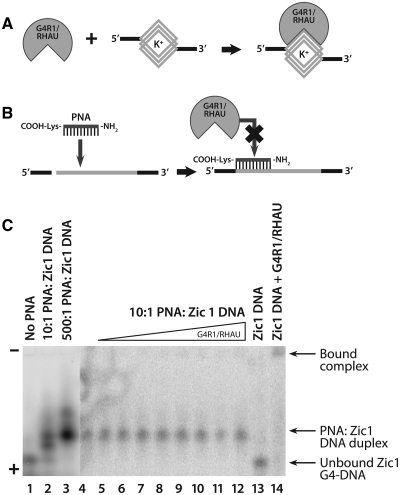
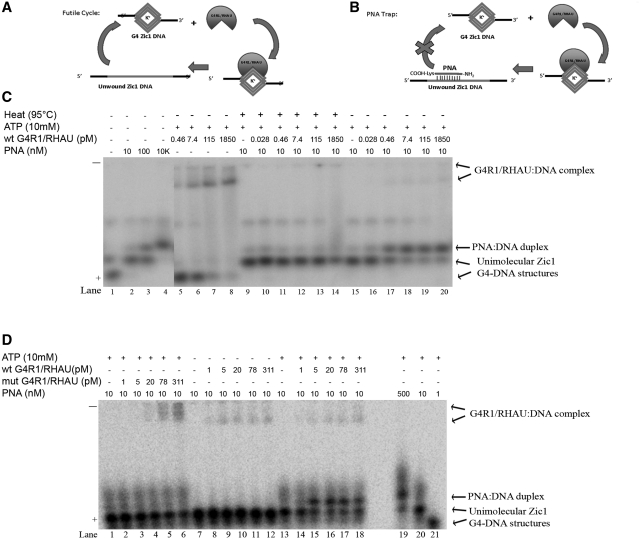
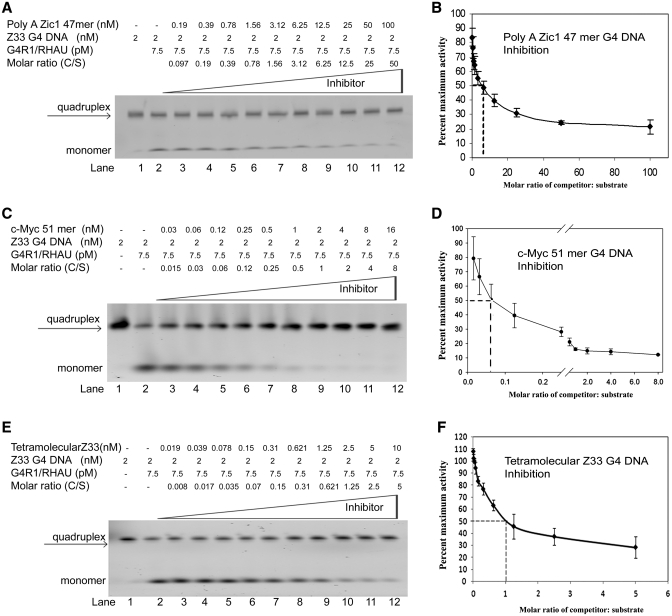
It has been previously shown that the DHX36 gene product, G4R1/RHAU, tightly binds tetramolecular G4-DNA with high affinity and resolves these structures into single strands. Here, we test the ability of G4R1/RHAU to bind and unwind unimolecular G4-DNA. Gel mobility shift assays were used to measure the binding affinity of G4R1/RHAU for unimolecular G4-DNA-formed sequences from the Zic1 gene and the c-Myc promoter. Extremely tight binding produced apparent K(d)'s of 6, 3 and 4 pM for two Zic1 G4-DNAs and a c-Myc G4-DNA, respectively. The low enzyme concentrations required for measuring these K(d)'s limit the precision of their determination to upper boundary estimates. Similar tight binding was not observed in control non-G4 forming DNA sequences or in single-stranded DNA having guanine-rich runs capable of forming tetramolecular G4-DNA. Using a peptide nucleic acid (PNA) trap assay, we show that G4R1/RHAU catalyzes unwinding of unimolecular Zic1 G4-DNA into an unstructured state capable of hybridizing to a complementary PNA. Binding was independent of adenosine triphosphate (ATP), but the PNA trap assay showed that unwinding of G4-DNA was ATP dependent. Competition studies indicated that unimolecular Zic1 and c-Myc G4-DNA structures inhibit G4R1/RHAU-catalyzed resolution of tetramolecular G4-DNA. This report provides evidence that G4R1/RHAU tightly binds and unwinds unimolecular G4-DNA structures.
Figures
References
-
- Mergny JL, Phan A-T, Lacroix L. Following G-quartet formation by UV-spectroscopy. FEBS Lett. 1998;435:74–78. - PubMed
-
- Sen D, Gilbert W. A sodium-potassium switch in the formation of four-stranded G4-DNA. Nature. 1990;344:410–414. - PubMed
-
- Prislan I, Lah J, Vesnaver G. Diverse polymorphism of G-quadruplexes as a kinetic phenomenon. J. Am. Chem. Soc. 2008;130:14161–14169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
