

Virulence differences in Toxoplasma mediated by amplification of a family of polymorphic pseudokinases
- PMID: 21586633
- PMCID: PMC3111276
- DOI: 10.1073/pnas.1015338108
Virulence differences in Toxoplasma mediated by amplification of a family of polymorphic pseudokinases
Abstract
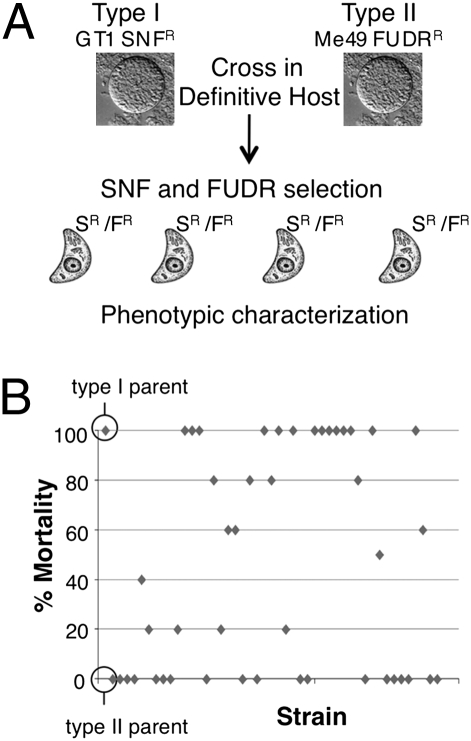
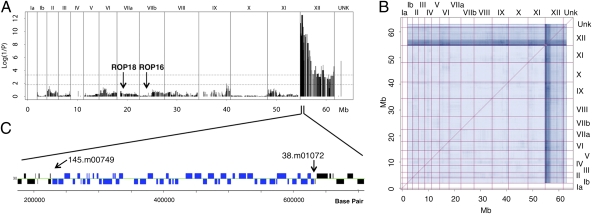
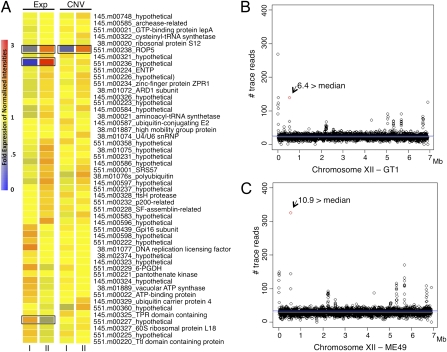
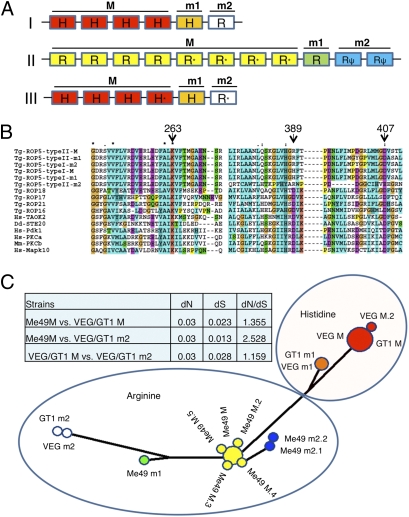
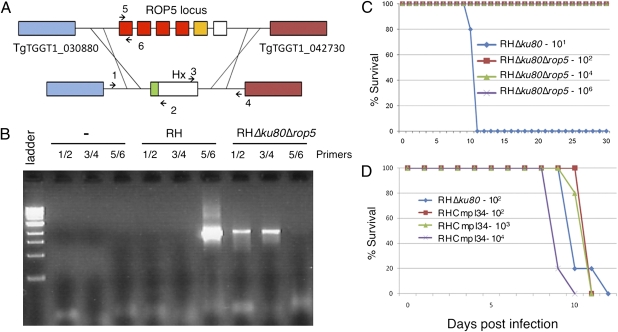
The population structure of Toxoplasma gondii includes three highly prevalent clonal lineages referred to as types I, II, and III, which differ greatly in virulence in the mouse model. Previous studies have implicated a family of serine/threonine protein kinases found in rhoptries (ROPs) as important in mediating virulence differences between strain types. Here, we explored the genetic basis of differences in virulence between the highly virulent type I lineage and moderately virulent type II based on successful genetic cross between these lineages. Genome-wide association revealed that a single quantitative trait locus controls the dramatic difference in lethality between these strain types. Neither ROP16 nor ROP18, previously implicated in virulence of T. gondii, was found to contribute to differences between types I and II. Instead, the major virulence locus contained a tandem cluster of polymorphic alleles of ROP5, which showed similar protein expression between strains. ROP5 contains a conserved serine/threonine protein kinase domain that includes only part of the catalytic triad, and hence, all members are considered to be pseudokinases. Genetic disruption of the entire ROP5 locus in the type I lineage led to complete attenuation of acute virulence, and complementation with ROP5 restored lethality to WT levels. These findings reveal that a locus of polymorphic pseudokinases plays an important role in pathogenesis of toxoplasmosis in the mouse model.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dubey JP. Toxoplasmosis of Animals and Humans. Boca Raton, FL: CRC; 2010.
-
- Joynson DH, Wreghitt TJ. Toxoplasmosis: A Comprehensive Clinical Guide. Cambridge, UK: Cambridge University Press; 2001.
-
- Jones LA, Alexander J, Roberts CW. Ocular toxoplasmosis: In the storm of the eye. Parasite Immunol. 2006;28:635–642. - PubMed
-
- Sibley LD, Ajioka JW. Population structure of Toxoplasma gondii: Clonal expansion driven by infrequent recombination and selective sweeps. Annu Rev Microbiol. 2008;62:329–351. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
