

A small zinc finger thylakoid protein plays a role in maintenance of photosystem II in Arabidopsis thaliana
- PMID: 21586683
- PMCID: PMC3123961
- DOI: 10.1105/tpc.111.085456
A small zinc finger thylakoid protein plays a role in maintenance of photosystem II in Arabidopsis thaliana
Abstract
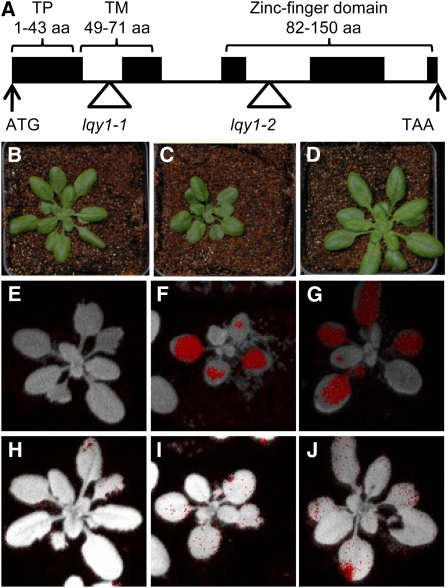
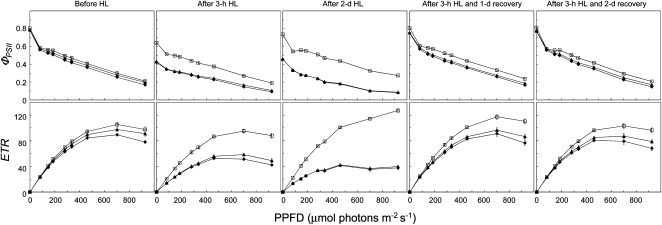
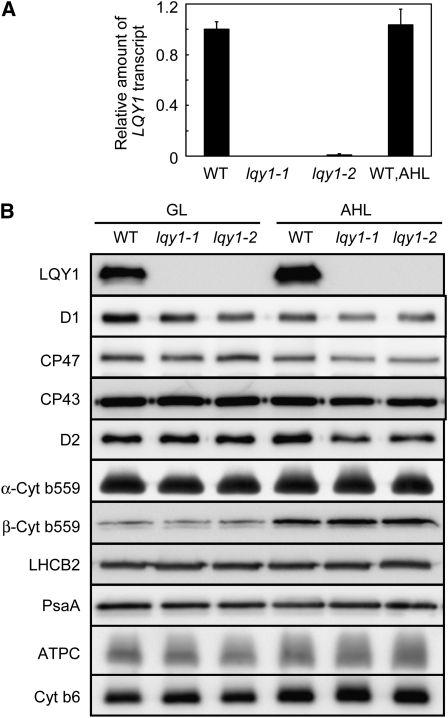
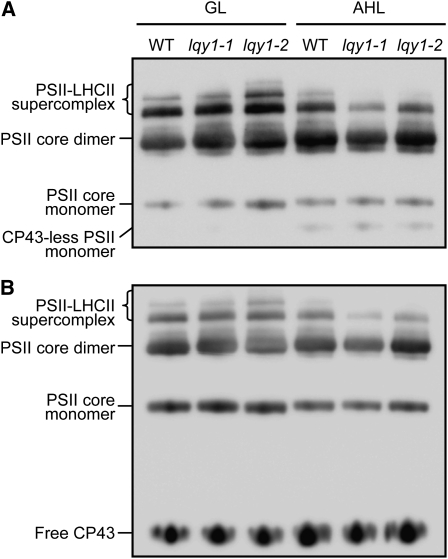
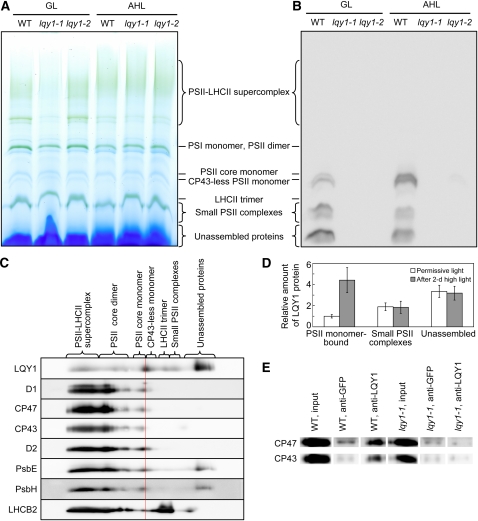
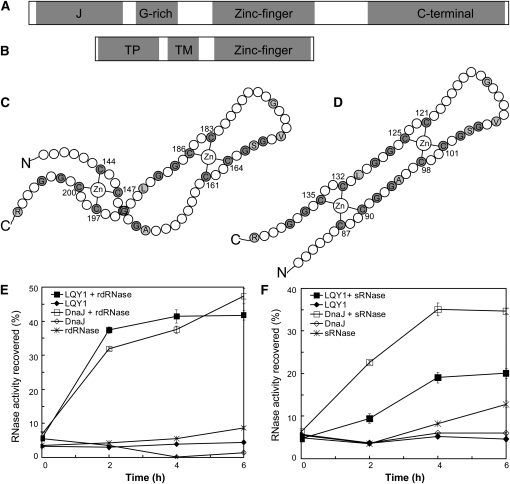
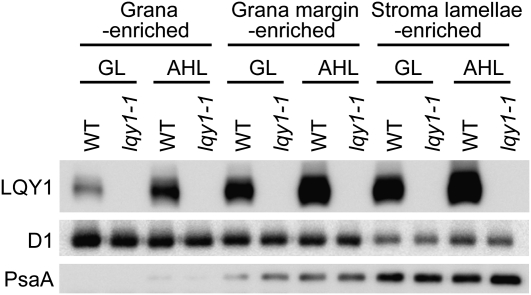
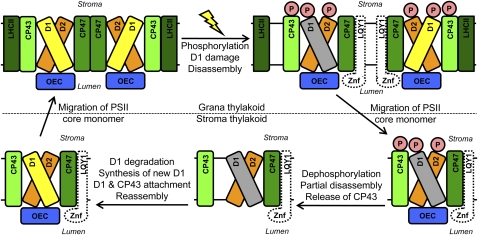
This work identifies LOW QUANTUM YIELD OF PHOTOSYSTEM II1 (LQY1), a Zn finger protein that shows disulfide isomerase activity, interacts with the photosystem II (PSII) core complex, and may act in repair of photodamaged PSII complexes. Two mutants of an unannotated small Zn finger containing a thylakoid membrane protein of Arabidopsis thaliana (At1g75690; LQY1) were found to have a lower quantum yield of PSII photochemistry and reduced PSII electron transport rate following high-light treatment. The mutants dissipate more excess excitation energy via nonphotochemical pathways than wild type, and they also display elevated accumulation of reactive oxygen species under high light. After high-light treatment, the mutants have less PSII-light-harvesting complex II supercomplex than wild-type plants. Analysis of thylakoid membrane protein complexes showed that wild-type LQY1 protein comigrates with the PSII core monomer and the CP43-less PSII monomer (a marker for ongoing PSII repair and reassembly). PSII repair and reassembly involve the breakage and formation of disulfide bonds among PSII proteins. Interestingly, the recombinant LQY1 protein demonstrates a protein disulfide isomerase activity. LQY1 is more abundant in stroma-exposed thylakoids, where key steps of PSII repair and reassembly take place. The absence of the LQY1 protein accelerates turnover and synthesis of PSII reaction center protein D1. These results suggest that the LQY1 protein may be involved in maintaining PSII activity under high light by regulating repair and reassembly of PSII complexes.
Figures

References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Aro E.-M., Suorsa M., Rokka A., Allahverdiyeva Y., Paakkarinen V., Saleem A., Battchikova N., Rintamäki E. (2005). Dynamics of photosystem II: A proteomic approach to thylakoid protein complexes. J. Exp. Bot. 56: 347–356 - PubMed
-
- Baker N.R., Harbinson J., Kramer D.M. (2007). Determining the limitations and regulation of photosynthetic energy transduction in leaves. Plant Cell Environ. 30: 1107–1125 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
