

Transcription factor IRF8 directs a silencing programme for TH17 cell differentiation
- PMID: 21587231
- PMCID: PMC3112536
- DOI: 10.1038/ncomms1311
Transcription factor IRF8 directs a silencing programme for TH17 cell differentiation
Abstract
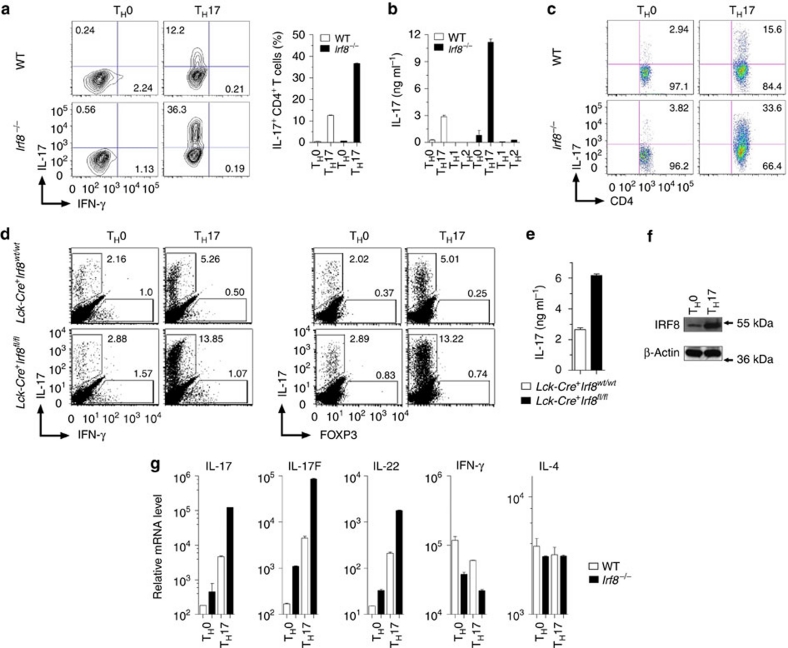
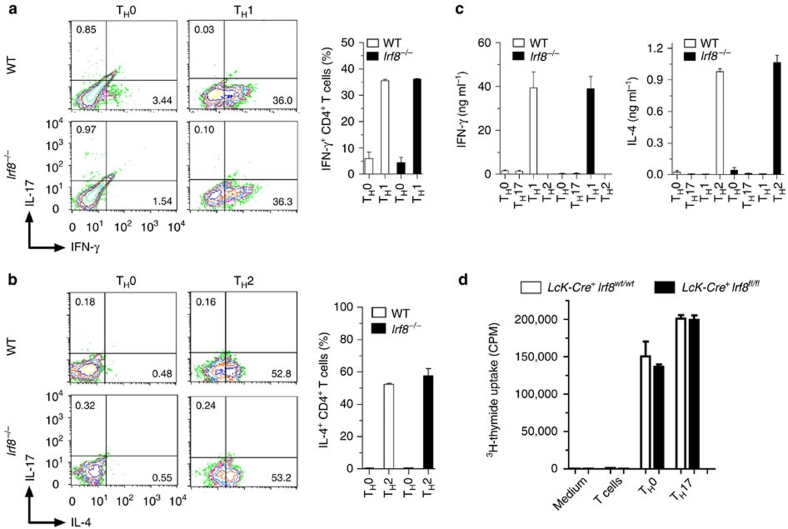
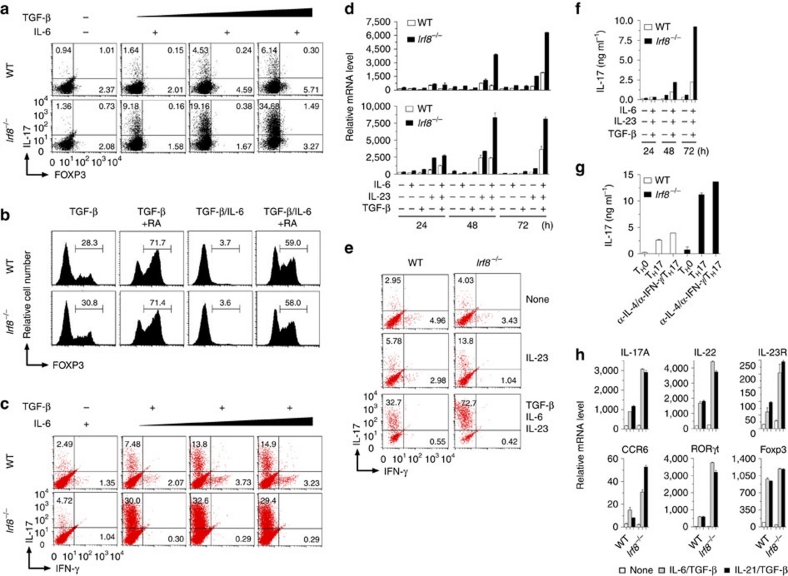
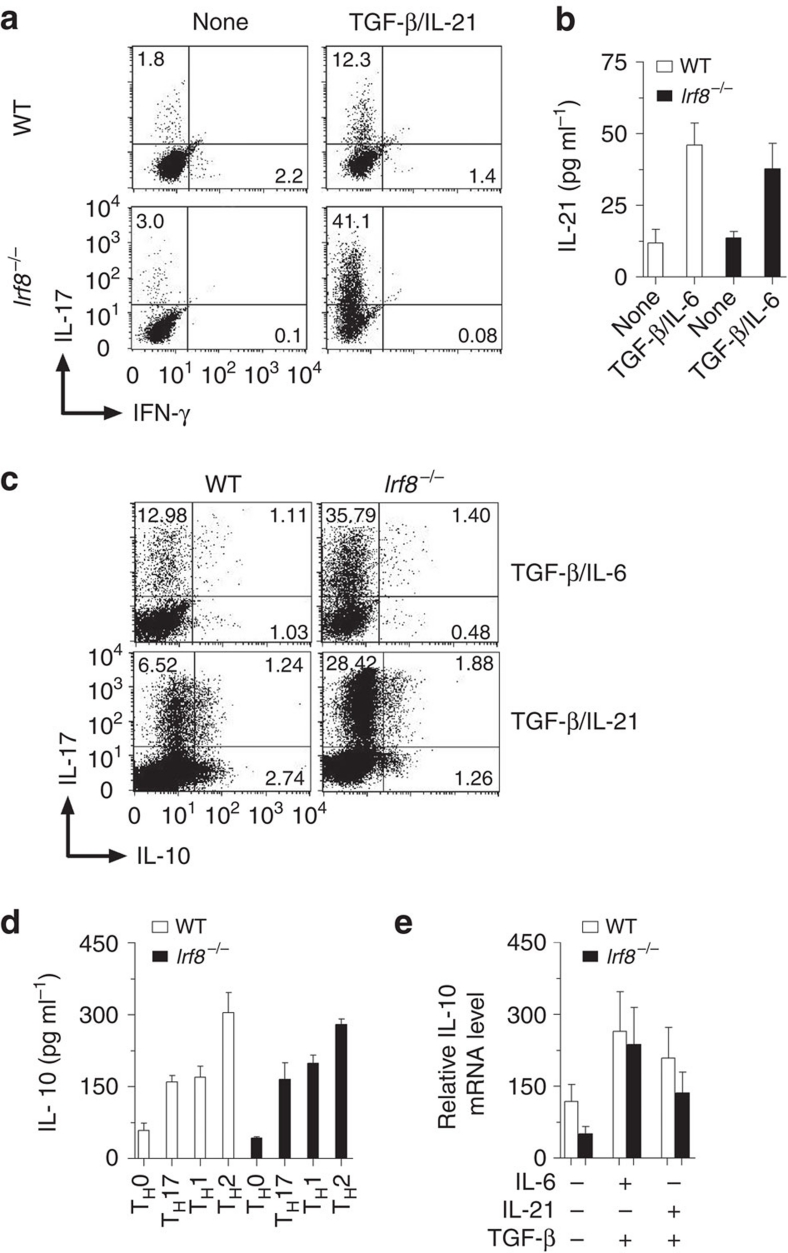
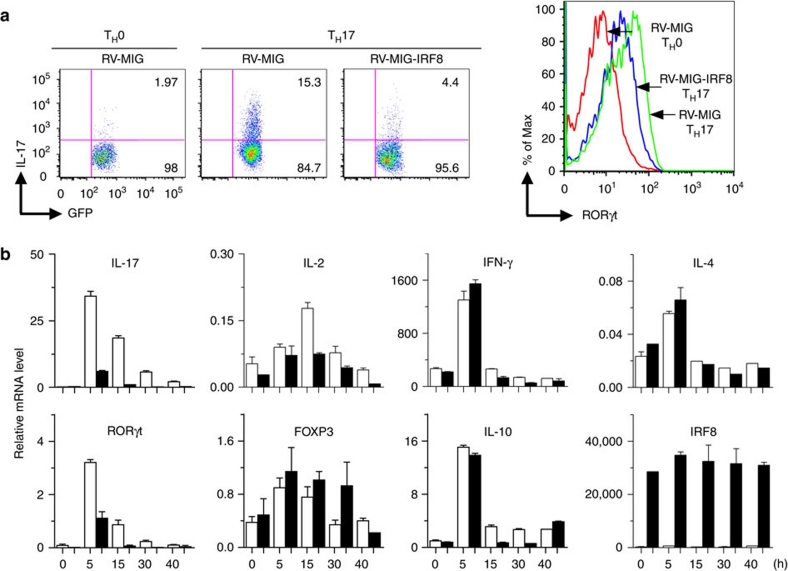
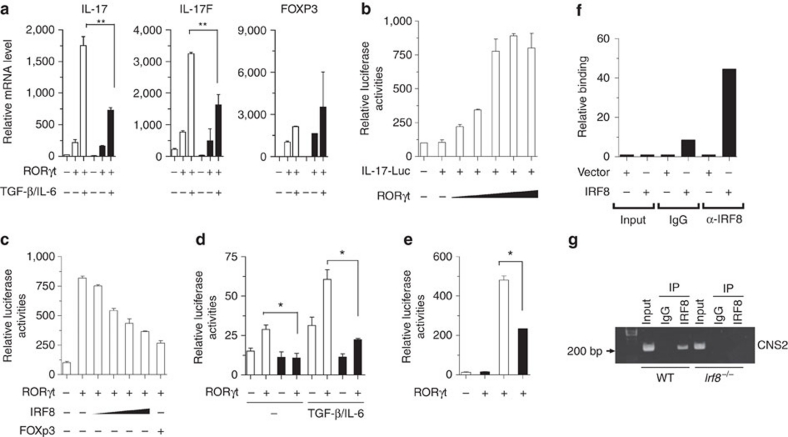
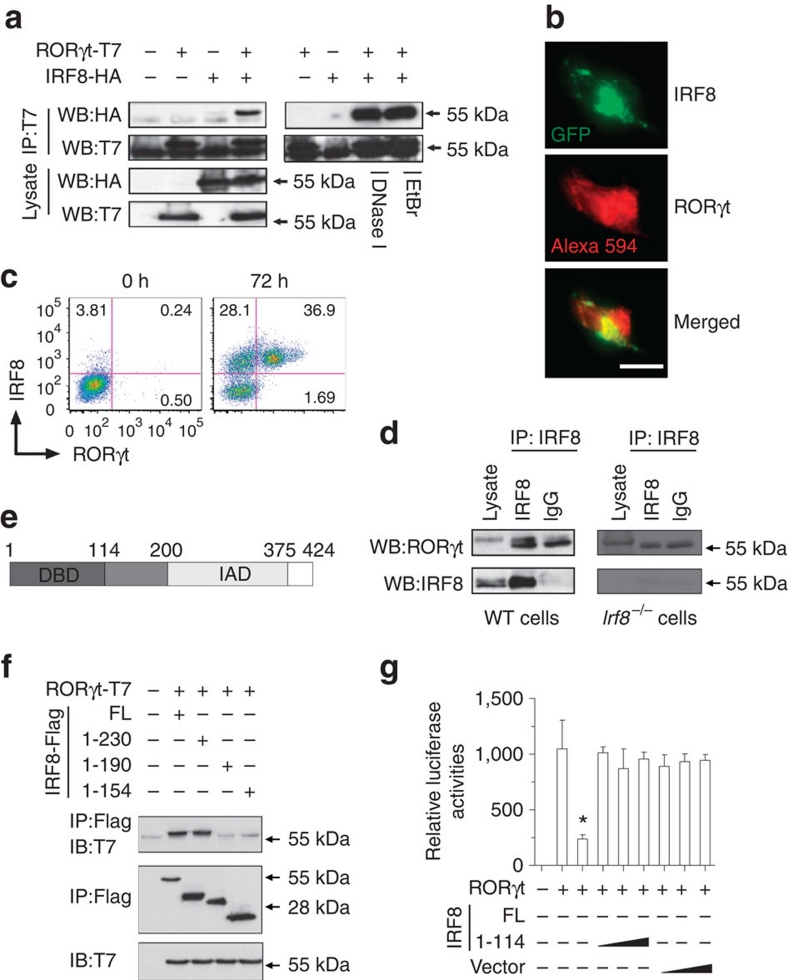
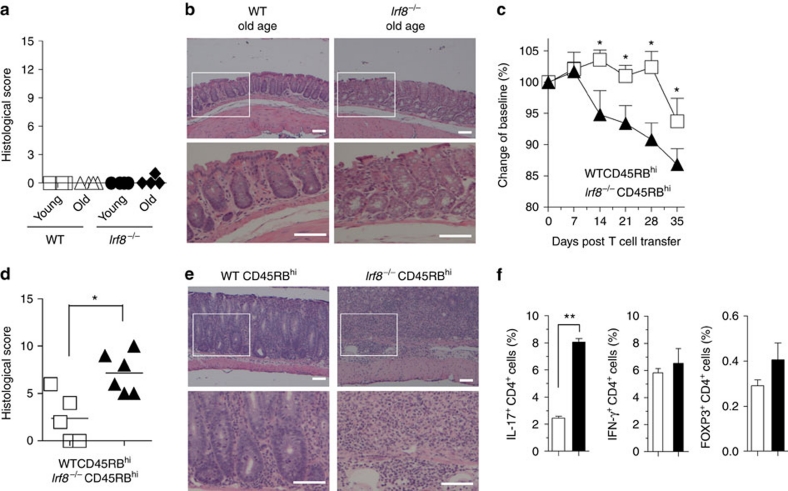
T(H)17 cells are recognized as a unique subset of T helper cells that have critical roles in the pathogenesis of autoimmunity and tissue inflammation. Although RORγt is necessary for the generation of T(H)17 cells, the molecular mechanisms underlying the functional diversity of T(H)17 cells are not fully understood. Here we show that a member of interferon regulatory factor (IRF) family of transcription factors, IRF8, has a critical role in silencing T(H)17-cell differentiation. Mice with a conventional knockout, as well as a T cell-specific deletion, of the Irf8 gene exhibited more efficient T(H)17 cells. Indeed, studies of an experimental model of colitis showed that IRF8 deficiency resulted in more severe inflammation with an enhanced T(H)17 phenotype. IRF8 was induced steadily and inhibited T(H)17-cell differentiation during T(H)17 lineage commitment at least in part through its physical interaction with RORγt. These findings define IRF8 as a novel intrinsic transcriptional inhibitor of T(H)17-cell differentiation.
Figures
References
-
- Weaver C. T., Harrington L. E., Mangan P. R., Gavrieli M. & Murphy K. M. Th17: an effector CD4 T cell lineage with regulatory T cell ties. Immunity 24, 677–688 (2006). - PubMed
-
- Zheng Y. et al. Interleukin-22, a T(H)17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature 445, 648–651 (2007). - PubMed
-
- Veldhoen M., Hocking R. J., Atkins C. J., Locksley R. M. & Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
