

Unc-51/ATG1 controls axonal and dendritic development via kinesin-mediated vesicle transport in the Drosophila brain
- PMID: 21589871
- PMCID: PMC3093397
- DOI: 10.1371/journal.pone.0019632
Unc-51/ATG1 controls axonal and dendritic development via kinesin-mediated vesicle transport in the Drosophila brain
Abstract
Background: Members of the evolutionary conserved Ser/Thr kinase Unc-51 family are key regulatory proteins that control neural development in both vertebrates and invertebrates. Previous studies have suggested diverse functions for the Unc-51 protein, including axonal elongation, growth cone guidance, and synaptic vesicle transport.
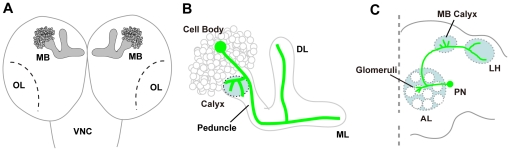
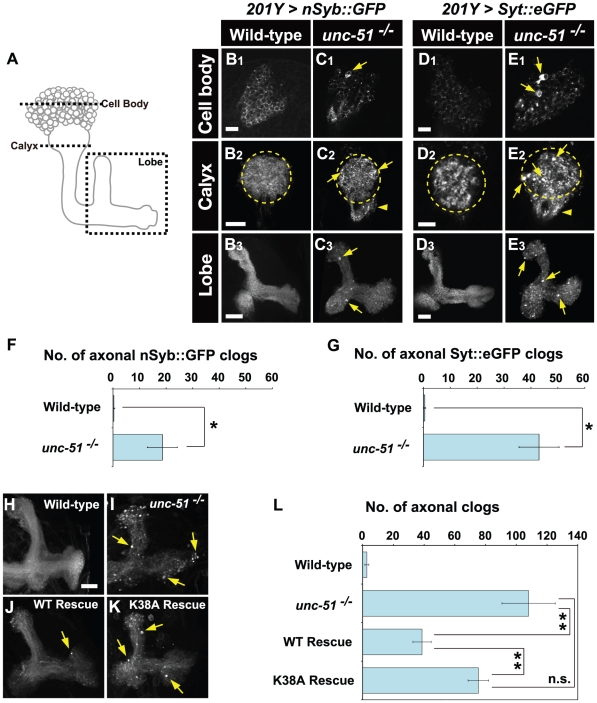
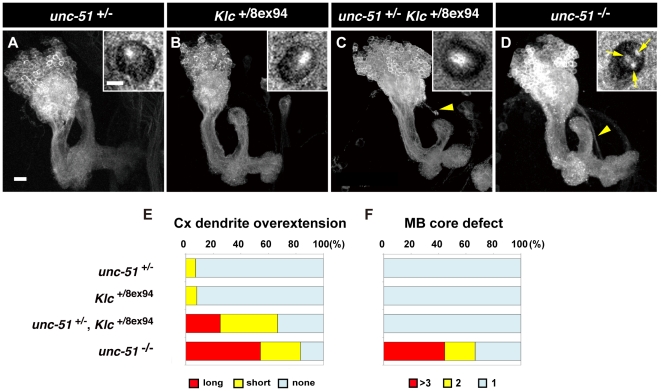
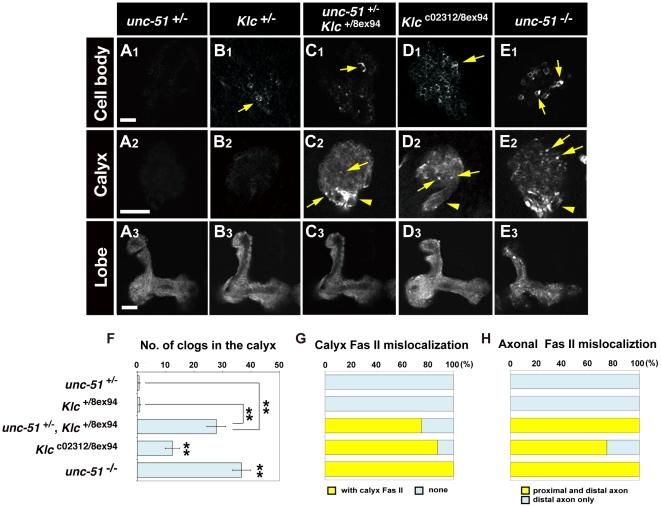
Methodology/principal findings: In this work, we have investigated the functional significance of Unc-51-mediated vesicle transport in the development of complex brain structures in Drosophila. We show that Unc-51 preferentially accumulates in newly elongating axons of the mushroom body, a center of olfactory learning in flies. Mutations in unc-51 cause disintegration of the core of the developing mushroom body, with mislocalization of Fasciclin II (Fas II), an IgG-family cell adhesion molecule important for axonal guidance and fasciculation. In unc-51 mutants, Fas II accumulates in the cell bodies, calyx, and the proximal peduncle. Furthermore, we show that mutations in unc-51 cause aberrant overshooting of dendrites in the mushroom body and the antennal lobe. Loss of unc-51 function leads to marked accumulation of Rab5 and Golgi components, whereas the localization of dendrite-specific proteins, such as Down syndrome cell adhesion molecule (DSCAM) and No distributive disjunction (Nod), remains unaltered. Genetic analyses of kinesin light chain (Klc) and unc-51 double heterozygotes suggest the importance of kinesin-mediated membrane transport for axonal and dendritic development. Moreover, our data demonstrate that loss of Klc activity causes similar axonal and dendritic defects in mushroom body neurons, recapitulating the salient feature of the developmental abnormalities caused by unc-51 mutations.
Conclusions/significance: Unc-51 plays pivotal roles in the axonal and dendritic development of the Drosophila brain. Unc-51-mediated membrane vesicle transport is important in targeted localization of guidance molecules and organelles that regulate elongation and compartmentalization of developing neurons.
Conflict of interest statement
Figures




Similar articles
-
UNC-51/ATG1 kinase regulates axonal transport by mediating motor-cargo assembly.Genes Dev. 2008 Dec 1;22(23):3292-307. doi: 10.1101/gad.1734608. Genes Dev. 2008. PMID: 19056884 Free PMC article.
-
The kinesin-3, unc-104 regulates dendrite morphogenesis and synaptic development in Drosophila.Genetics. 2013 Sep;195(1):59-72. doi: 10.1534/genetics.113.151639. Epub 2013 Jun 14. Genetics. 2013. PMID: 23770702 Free PMC article.
-
The kinesin-associated protein UNC-76 is required for axonal transport in the Drosophila nervous system.Mol Biol Cell. 2003 Aug;14(8):3356-65. doi: 10.1091/mbc.e02-12-0800. Epub 2003 May 3. Mol Biol Cell. 2003. PMID: 12925768 Free PMC article.
-
Smart motors and cargo steering drive kinesin-mediated selective transport.Mol Cell Neurosci. 2020 Mar;103:103464. doi: 10.1016/j.mcn.2019.103464. Epub 2020 Jan 20. Mol Cell Neurosci. 2020. PMID: 31972342 Free PMC article. Review.
-
The importance of fasciculation and elongation protein zeta-1 in neural circuit establishment and neurological disorders.Neural Regen Res. 2022 Jun;17(6):1165-1171. doi: 10.4103/1673-5374.327327. Neural Regen Res. 2022. PMID: 34782550 Free PMC article. Review.
Cited by
-
Monensin Sensitive 1 Regulates Dendritic Arborization in Drosophila by Modulating Endocytic Flux.Front Cell Dev Biol. 2019 Aug 2;7:145. doi: 10.3389/fcell.2019.00145. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31428611 Free PMC article.
-
The PARK10 gene USP24 is a negative regulator of autophagy and ULK1 protein stability.Autophagy. 2020 Jan;16(1):140-153. doi: 10.1080/15548627.2019.1598754. Epub 2019 Apr 7. Autophagy. 2020. PMID: 30957634 Free PMC article.
-
The Atg1-Tor pathway regulates yolk catabolism in Drosophila embryos.Development. 2015 Nov 15;142(22):3869-78. doi: 10.1242/dev.125419. Epub 2015 Sep 22. Development. 2015. PMID: 26395483 Free PMC article.
-
Atg13 Is Essential for Autophagy and Cardiac Development in Mice.Mol Cell Biol. 2015 Dec 7;36(4):585-95. doi: 10.1128/MCB.01005-15. Print 2016 Feb 15. Mol Cell Biol. 2015. PMID: 26644405 Free PMC article.
-
The ALS-FTD-linked gene product, C9orf72, regulates neuronal morphogenesis via autophagy.Autophagy. 2019 May;15(5):827-842. doi: 10.1080/15548627.2019.1569441. Epub 2019 Jan 28. Autophagy. 2019. PMID: 30669939 Free PMC article.
References
-
- Guzik BW, Goldstein LS. Microtubule-dependent transport in neurons: steps towards an understanding of regulation, function and dysfunction. Curr Opin Cell Biol. 2004;16:443–450. - PubMed
-
- Sann S, Wang Z, Brown H, Jin Y. Roles of endosomal trafficking in neurite outgrowth and guidance. Trends Cell Biol. 2009;19:317–324. - PubMed
-
- Hirokawa N, Takemura R. Molecular motors in neuronal development, intracellular transport and diseases. Curr Opin Neurobiol. 2004;14:564–573. - PubMed
-
- Horton AC, Ehlers MD. Neuronal polarity and trafficking. Neuron. 2003;40:277–295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
