

An E2F1-mediated DNA damage response contributes to the replication of human cytomegalovirus
- PMID: 21589897
- PMCID: PMC3093362
- DOI: 10.1371/journal.ppat.1001342
An E2F1-mediated DNA damage response contributes to the replication of human cytomegalovirus
Abstract
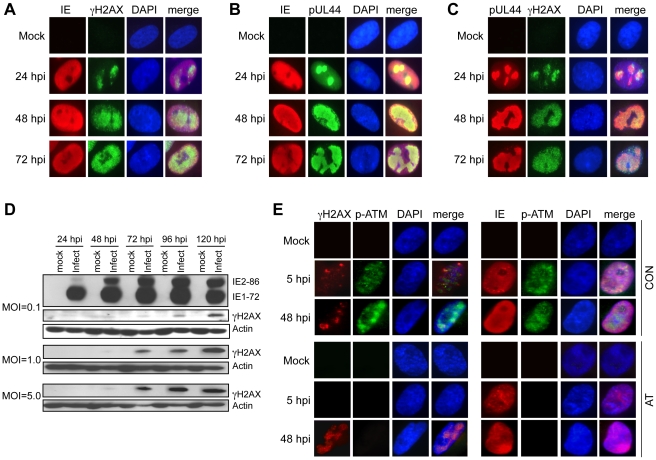
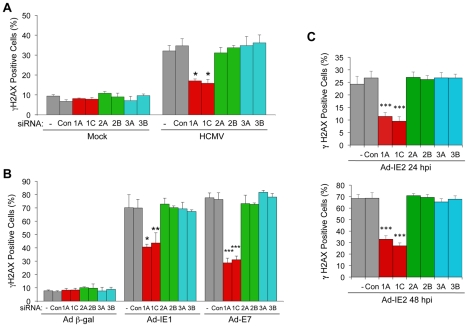
DNA damage resulting from intrinsic or extrinsic sources activates DNA damage responses (DDRs) centered on protein kinase signaling cascades. The usual consequences of inducing DDRs include the activation of cell cycle checkpoints together with repair of the damaged DNA or induction of apoptosis. Many DNA viruses elicit host DDRs during infection and some viruses require the DDR for efficient replication. However, the mechanism by which DDRs are activated by viral infection is poorly understood. Human cytomegalovirus (HCMV) infection induces a DDR centered on the activation of ataxia telangiectasia mutated (ATM) protein kinase. Here we show that HCMV replication is compromised in cells with inactivated or depleted ATM and that ATM is essential for the host DDR early during infection. Likewise, a downstream target of ATM phosphorylation, H2AX, also contributes to viral replication. The ATM-dependent DDR is detected as discrete, nuclear γH2AX foci early in infection and can be activated by IE proteins. By 24 hpi, γH2AX is observed primarily in HCMV DNA replication compartments. We identified a role for the E2F1 transcription factor in mediating this DDR and viral replication. E2F1, but not E2F2 or E2F3, promotes the accumulation of γH2AX during HCMV infection or IE protein expression. Moreover, E2F1 expression, but not the expression of E2F2 or E2F3, is required for efficient HCMV replication. These results reveal a novel role for E2F1 in mediating an ATM-dependent DDR that contributes to viral replication. Given that E2F activity is often deregulated by infection with DNA viruses, these observations raise the possibility that an E2F1-mediated mechanism of DDR activation may be conserved among DNA viruses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
Human cytomegalovirus disrupts both ataxia telangiectasia mutated protein (ATM)- and ATM-Rad3-related kinase-mediated DNA damage responses during lytic infection.J Virol. 2007 Feb;81(4):1934-50. doi: 10.1128/JVI.01670-06. Epub 2006 Dec 6. J Virol. 2007. PMID: 17151099 Free PMC article.
-
Loss of the tumor suppressor BIN1 enables ATM Ser/Thr kinase activation by the nuclear protein E2F1 and renders cancer cells resistant to cisplatin.J Biol Chem. 2019 Apr 5;294(14):5700-5719. doi: 10.1074/jbc.RA118.005699. Epub 2019 Feb 7. J Biol Chem. 2019. PMID: 30733337 Free PMC article.
-
DNA Damage Signaling Is Induced in the Absence of Epstein-Barr Virus (EBV) Lytic DNA Replication and in Response to Expression of ZEBRA.PLoS One. 2015 May 7;10(5):e0126088. doi: 10.1371/journal.pone.0126088. eCollection 2015. PLoS One. 2015. PMID: 25950714 Free PMC article.
-
DNA-damage response pathways triggered by viral replication.Expert Rev Mol Med. 2006 Mar 3;8(5):1-11. doi: 10.1017/S1462399406010544. Expert Rev Mol Med. 2006. PMID: 16515730 Free PMC article. Review.
-
The NEMO/ATM/IKKα complex: A key player in colorectal cancer progression and treatment.Sci Signal. 2025 Mar 11;18(877):eadv1265. doi: 10.1126/scisignal.adv1265. Epub 2025 Mar 11. Sci Signal. 2025. PMID: 40067906 Review.
Cited by
-
The Transcription and Translation Landscapes during Human Cytomegalovirus Infection Reveal Novel Host-Pathogen Interactions.PLoS Pathog. 2015 Nov 24;11(11):e1005288. doi: 10.1371/journal.ppat.1005288. eCollection 2015. PLoS Pathog. 2015. PMID: 26599541 Free PMC article.
-
Human cytomegalovirus-induced host protein citrullination is crucial for viral replication.Nat Commun. 2021 Jun 23;12(1):3910. doi: 10.1038/s41467-021-24178-6. Nat Commun. 2021. PMID: 34162877 Free PMC article.
-
Complex roles for proliferating cell nuclear antigen in restricting human cytomegalovirus replication.bioRxiv [Preprint]. 2025 Jan 6:2025.01.06.631530. doi: 10.1101/2025.01.06.631530. bioRxiv. 2025. Update in: mBio. 2025 May 14;16(5):e0045025. doi: 10.1128/mbio.00450-25. PMID: 39829879 Free PMC article. Updated. Preprint.
-
E2F1 suppresses Epstein-Barr virus lytic reactivation through cellular and viral transcriptional networks.PLoS Pathog. 2025 Aug 7;21(8):e1013410. doi: 10.1371/journal.ppat.1013410. eCollection 2025 Aug. PLoS Pathog. 2025. PMID: 40773523 Free PMC article.
-
ATM supports gammaherpesvirus replication by attenuating type I interferon pathway.Virology. 2017 Oct;510:137-146. doi: 10.1016/j.virol.2017.07.014. Epub 2017 Jul 18. Virology. 2017. PMID: 28732227 Free PMC article.
References
-
- Bartek J, Lukas J. DNA damage checkpoints: from initiation to recovery or adaptation. Curr Opin Cell Biol. 2007;19:238–245. - PubMed
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
-
- Lau A, Swinbank KM, Ahmed PS, Taylor DL, Jackson SP, et al. Suppression of HIV-1 infection by a small molecule inhibitor of the ATM kinase. Nat Cell Biol. 2005;7:493–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
