

A family of helminth molecules that modulate innate cell responses via molecular mimicry of host antimicrobial peptides
- PMID: 21589904
- PMCID: PMC3093369
- DOI: 10.1371/journal.ppat.1002042
A family of helminth molecules that modulate innate cell responses via molecular mimicry of host antimicrobial peptides
Abstract
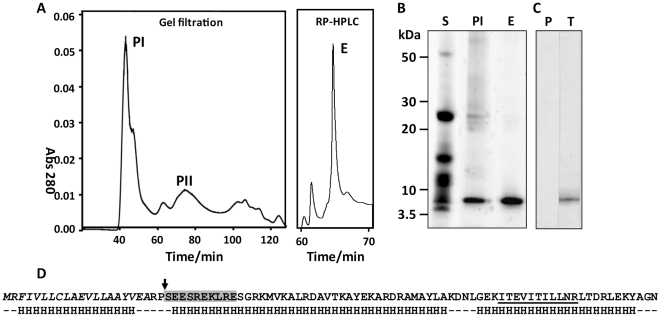
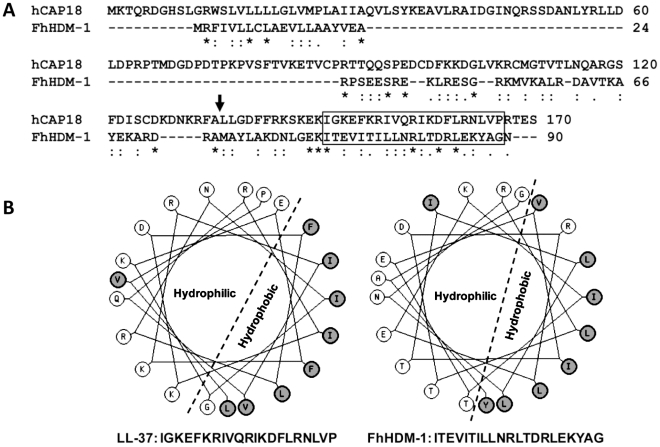
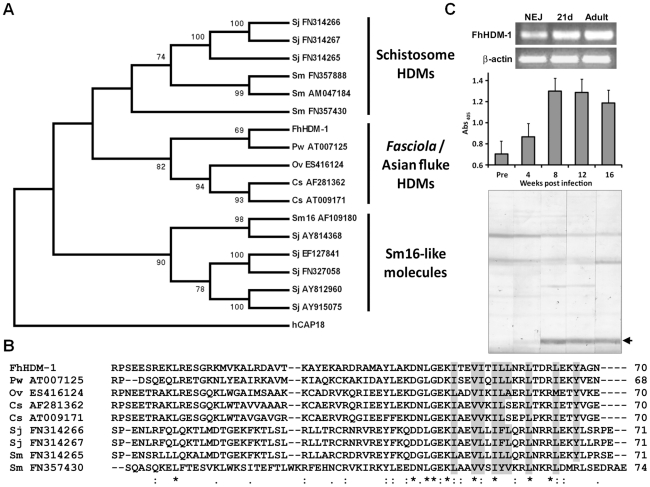
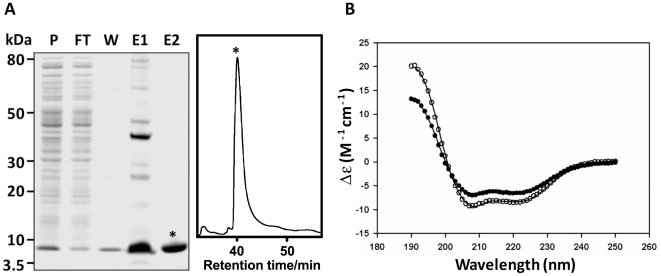
Over the last decade a significant number of studies have highlighted the central role of host antimicrobial (or defence) peptides in modulating the response of innate immune cells to pathogen-associated ligands. In humans, the most widely studied antimicrobial peptide is LL-37, a 37-residue peptide containing an amphipathic helix that is released via proteolytic cleavage of the precursor protein CAP18. Owing to its ability to protect against lethal endotoxaemia and clinically-relevant bacterial infections, LL-37 and its derivatives are seen as attractive candidates for anti-sepsis therapies. We have identified a novel family of molecules secreted by parasitic helminths (helminth defence molecules; HDMs) that exhibit similar biochemical and functional characteristics to human defence peptides, particularly CAP18. The HDM secreted by Fasciola hepatica (FhHDM-1) adopts a predominantly α-helical structure in solution. Processing of FhHDM-1 by F. hepatica cathepsin L1 releases a 34-residue C-terminal fragment containing a conserved amphipathic helix. This is analogous to the proteolytic processing of CAP18 to release LL-37, which modulates innate cell activation by classical toll-like receptor (TLR) ligands such as lipopolysaccharide (LPS). We show that full-length recombinant FhHDM-1 and a peptide analogue of the amphipathic C-terminus bind directly to LPS in a concentration-dependent manner, reducing its interaction with both LPS-binding protein (LBP) and the surface of macrophages. Furthermore, FhHDM-1 and the amphipathic C-terminal peptide protect mice against LPS-induced inflammation by significantly reducing the release of inflammatory mediators from macrophages. We propose that HDMs, by mimicking the function of host defence peptides, represent a novel family of innate cell modulators with therapeutic potential in anti-sepsis treatments and prevention of inflammation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A helminth cathelicidin-like protein suppresses antigen processing and presentation in macrophages via inhibition of lysosomal vATPase.FASEB J. 2012 Nov;26(11):4614-27. doi: 10.1096/fj.12-213876. Epub 2012 Aug 7. FASEB J. 2012. PMID: 22872675
-
An immunoregulatory amphipathic peptide derived from Fasciola hepatica helminth defense molecule (FhHDM-1.C2) exhibits potent biotherapeutic activity in a murine model of multiple sclerosis.FASEB J. 2025 Feb 28;39(4):e70380. doi: 10.1096/fj.202400793RR. FASEB J. 2025. PMID: 39950309 Free PMC article.
-
The immune modulatory peptide FhHDM-1 secreted by the helminth Fasciola hepatica prevents NLRP3 inflammasome activation by inhibiting endolysosomal acidification in macrophages.FASEB J. 2017 Jan;31(1):85-95. doi: 10.1096/fj.201500093R. Epub 2016 Sep 28. FASEB J. 2017. PMID: 27682204
-
Fasciola hepatica-Derived Molecules as Regulators of the Host Immune Response.Front Immunol. 2020 Sep 2;11:2182. doi: 10.3389/fimmu.2020.02182. eCollection 2020. Front Immunol. 2020. PMID: 32983184 Free PMC article. Review.
-
A parasitic helminth-derived peptide that targets the macrophage lysosome is a novel therapeutic option for autoimmune disease.Immunobiology. 2015 Feb;220(2):262-9. doi: 10.1016/j.imbio.2014.11.008. Epub 2014 Nov 18. Immunobiology. 2015. PMID: 25466586 Review.
Cited by
-
Lectin-Mediated Bacterial Modulation by the Intestinal Nematode Ascaris suum.Int J Mol Sci. 2021 Aug 14;22(16):8739. doi: 10.3390/ijms22168739. Int J Mol Sci. 2021. PMID: 34445445 Free PMC article.
-
The Extracellular Vesicles of the Helminth Pathogen, Fasciola hepatica: Biogenesis Pathways and Cargo Molecules Involved in Parasite Pathogenesis.Mol Cell Proteomics. 2015 Dec;14(12):3258-73. doi: 10.1074/mcp.M115.053934. Epub 2015 Oct 20. Mol Cell Proteomics. 2015. PMID: 26486420 Free PMC article.
-
Functional Ultrastructure of the Excretory Gland Cell in Zoonotic Anisakids (Anisakidae, Nematoda).Cells. 2019 Nov 17;8(11):1451. doi: 10.3390/cells8111451. Cells. 2019. PMID: 31744245 Free PMC article.
-
Type I Cystatin Derived from Fasciola gigantica Suppresses Macrophage-Mediated Inflammatory Responses.Pathogens. 2023 Mar 1;12(3):395. doi: 10.3390/pathogens12030395. Pathogens. 2023. PMID: 36986318 Free PMC article.
-
Complementary transcriptomic and proteomic analyses reveal the cellular and molecular processes that drive growth and development of Fasciola hepatica in the host liver.BMC Genomics. 2021 Jan 11;22(1):46. doi: 10.1186/s12864-020-07326-y. BMC Genomics. 2021. PMID: 33430759 Free PMC article.
References
-
- Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449:819–826. - PubMed
-
- Castellheim A, Brekke OL, Espevik T, Harboe M, Mollnes TE. Innate immune responses to danger signals in systemic inflammatory response syndrome and sepsis. Scand J Immunol. 2009;69:479–491. - PubMed
-
- Giuliani A, Pirri G, Rinaldi AC. Antimicrobial peptides: the LPS connection. Methods Mol Biol. 2010;618:137–154. - PubMed
-
- Hirsch T, Metzig M, Niederbichler A, Steinau HU, Eriksson E, et al. Role of host defense peptides of the innate immune response in sepsis. Shock. 2008;30:117–126. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
