

WRKY transcription factors involved in activation of SA biosynthesis genes
- PMID: 21595875
- PMCID: PMC3120740
- DOI: 10.1186/1471-2229-11-89
WRKY transcription factors involved in activation of SA biosynthesis genes
Abstract
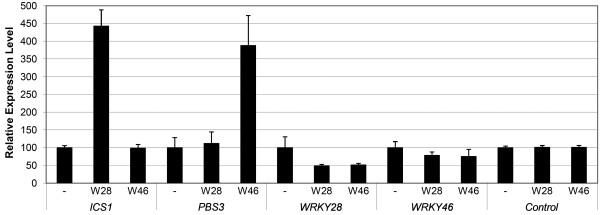
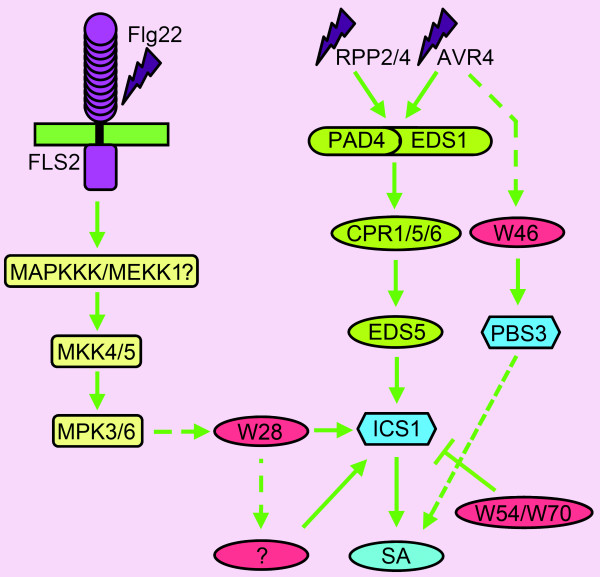
Background: Increased defense against a variety of pathogens in plants is achieved through activation of a mechanism known as systemic acquired resistance (SAR). The broad-spectrum resistance brought about by SAR is mediated through salicylic acid (SA). An important step in SA biosynthesis in Arabidopsis is the conversion of chorismate to isochorismate through the action of isochorismate synthase, encoded by the ICS1 gene. Also AVRPPHB SUSCEPTIBLE 3 (PBS3) plays an important role in SA metabolism, as pbs3 mutants accumulate drastically reduced levels of SA-glucoside, a putative storage form of SA. Bioinformatics analysis previously performed by us identified WRKY28 and WRKY46 as possible regulators of ICS1 and PBS3.
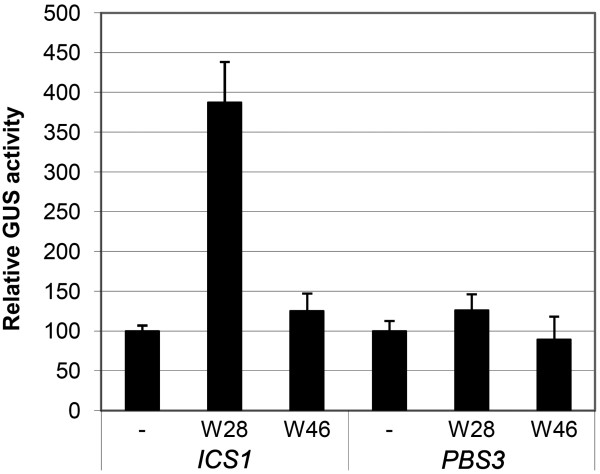
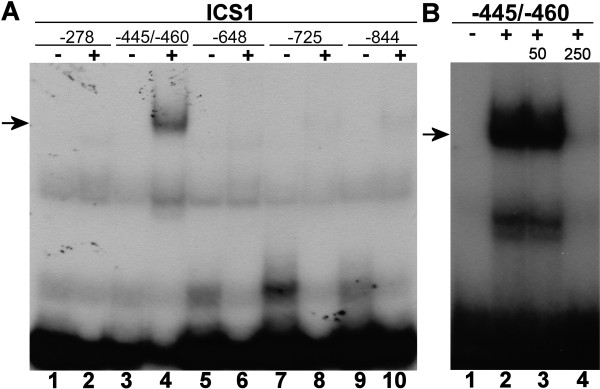
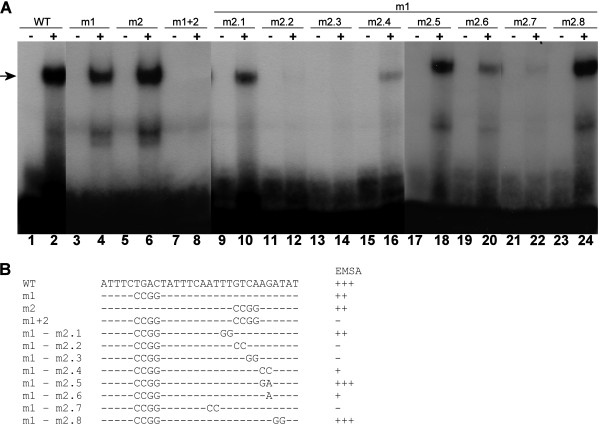
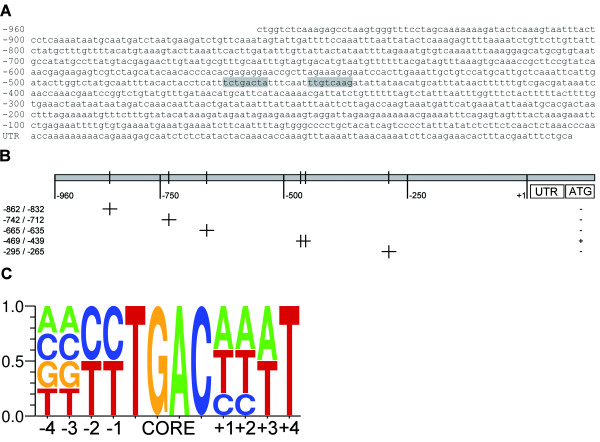
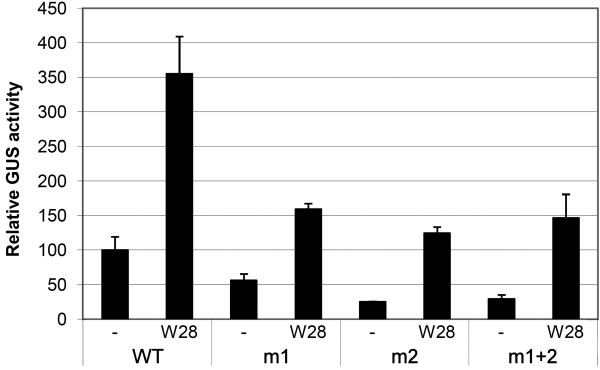
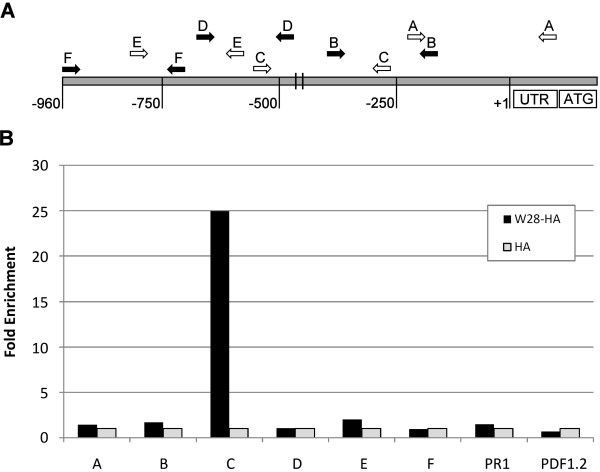
Results: Expression studies with ICS1 promoter::β-glucuronidase (GUS) genes in Arabidopsis thaliana protoplasts cotransfected with 35S::WRKY28 showed that over expression of WRKY28 resulted in a strong increase in GUS expression. Moreover, qRT-PCR analyses indicated that the endogenous ICS1 and PBS3 genes were highly expressed in protoplasts overexpressing WRKY28 or WRKY46, respectively. Electrophoretic mobility shift assays indentified potential WRKY28 binding sites in the ICS1 promoter, positioned -445 and -460 base pairs upstream of the transcription start site. Mutation of these sites in protoplast transactivation assays showed that these binding sites are functionally important for activation of the ICS1 promoter. Chromatin immunoprecipitation assays with haemagglutinin-epitope-tagged WRKY28 showed that the region of the ICS1 promoter containing the binding sites at -445 and -460 was highly enriched in the immunoprecipitated DNA.
Conclusions: The results obtained here confirm results from our multiple microarray co-expression analyses indicating that WRKY28 and WRKY46 are transcriptional activators of ICS1 and PBS3, respectively, and support this in silico screening as a powerful tool for identifying new components of stress signaling pathways.
Figures
References
-
- Métraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M, Gaudin J, Raschdorf K, Schmid E, Blum W, Inverardi B. Increase in Salicylic Acid at the Onset of Systemic Acquired Resistance in Cucumber. Science. 1990;250:1004–1006. - PubMed
-
- Malamy J, Carr JP, Klessig DF, Raskin I. Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science. 1990;250:1002–1004. - PubMed
-
- Dempsey DMA, Shah J, Klessig DF. Salicylic acid and disease resistance in plants. Crit Rev Plant Sci. 1999;18:547–575.
-
- Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annual Review of Phytopathology. 2005;43:205–227. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
