

The evolutionary history of the stearoyl-CoA desaturase gene family in vertebrates
- PMID: 21595943
- PMCID: PMC3112091
- DOI: 10.1186/1471-2148-11-132
The evolutionary history of the stearoyl-CoA desaturase gene family in vertebrates
Abstract
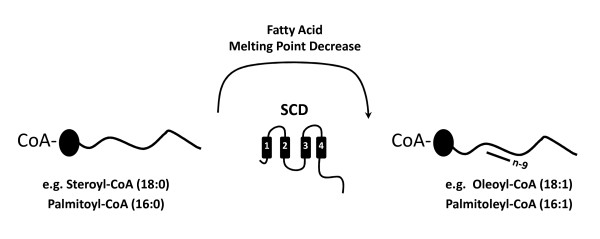
Background: Stearoyl-CoA desaturases (SCDs) are key enzymes involved in de novo monounsaturated fatty acid synthesis. They catalyze the desaturation of saturated fatty acyl-CoA substrates at the delta-9 position, generating essential components of phospholipids, triglycerides, cholesterol esters and wax esters. Despite being crucial for interpreting SCDs roles across species, the evolutionary history of the SCD gene family in vertebrates has yet to be elucidated, in particular their isoform diversity, origin and function. This work aims to contribute to this fundamental effort.
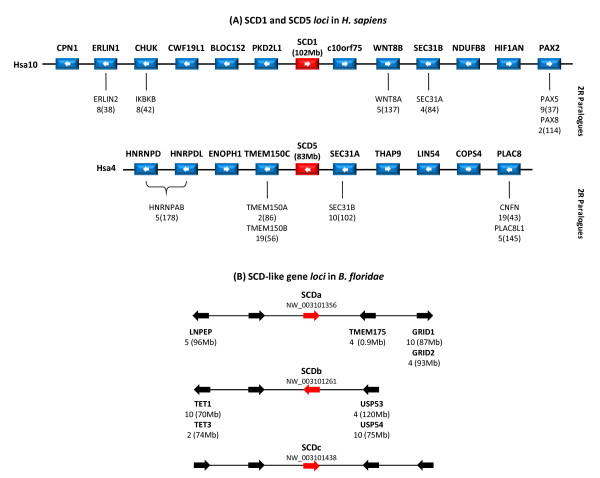
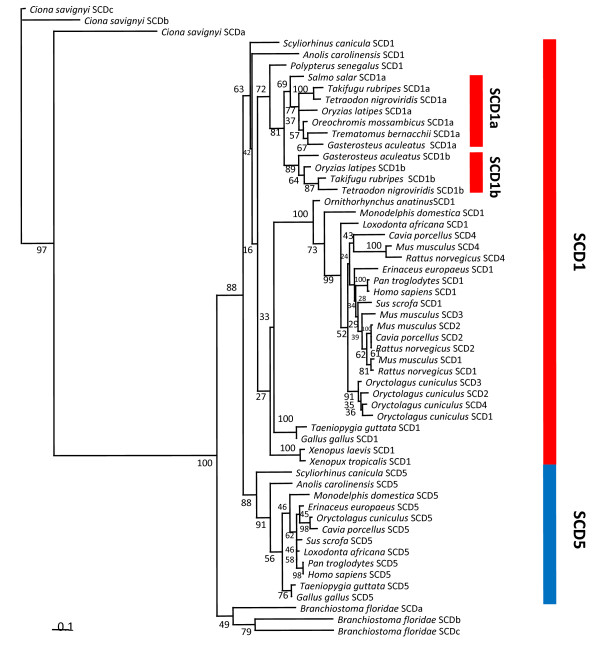
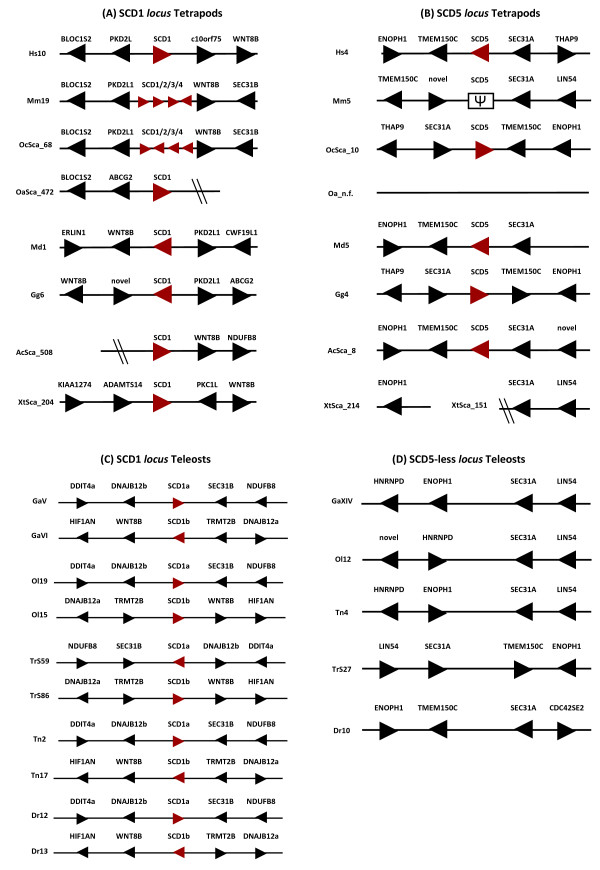
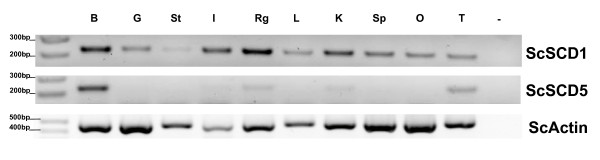
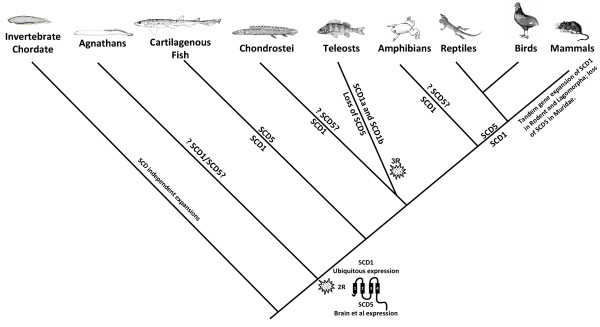
Results: We show here, through comparative genomics and phylogenetics that the SCD gene family underwent an unexpectedly complex history of duplication and loss events. Paralogy analysis hints that SCD1 and SCD5 genes emerged as part of the whole genome duplications (2R) that occurred at the stem of the vertebrate lineage. The SCD1 gene family expanded in rodents with the parallel loss of SCD5 in the Muridae family. The SCD1 gene expansion is also observed in the Lagomorpha although without the SCD5 loss. In the amphibian Xenopus tropicalis we find a single SCD1 gene but not SCD5, though this could be due to genome incompleteness. In the analysed teleost species no SCD5 is found, while the surrounding SCD5-less locus is conserved in comparison to tetrapods. In addition, the teleost SCD1 gene repertoire expanded to two copies as a result of the teleost specific genome duplication (3R). Finally, we describe clear orthologues of SCD1 and SCD5 in the chondrichthian, Scyliorhinus canicula, a representative of the oldest extant jawed vertebrate clade. Expression analysis in S. canicula shows that whilst SCD1 is ubiquitous, SCD5 is mainly expressed in the brain, a pattern which might indicate an evolutionary conserved function.
Conclusion: We conclude that the SCD1 and SCD5 genes emerged as part of the 2R genome duplications. We propose that the evolutionary conserved gene expression between distinct lineages underpins the importance of SCD activity in the brain (and probably the pancreas), in a yet to be defined role. We argue that an expression independent of an external stimulus, such as diet induced activity, emerged as a novel function in vertebrate ancestry allocated to the SCD5 isoform in various tissues (e.g. brain and pancreas), and it was selectively maintained throughout vertebrate evolution.
Figures
References
-
- Enoch HG, Catalá A, Strittmatter P. Mechanism of rat liver microsomal stearyl-CoA desaturase. Studies of the substrate specificity, enzyme-substrate interactions, and the function of lipid. J Biol Chem. 1976;251(16):5095–103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
