

Poly(ADP-ribose) regulates stress responses and microRNA activity in the cytoplasm
- PMID: 21596313
- PMCID: PMC3898460
- DOI: 10.1016/j.molcel.2011.04.015
Poly(ADP-ribose) regulates stress responses and microRNA activity in the cytoplasm
Abstract
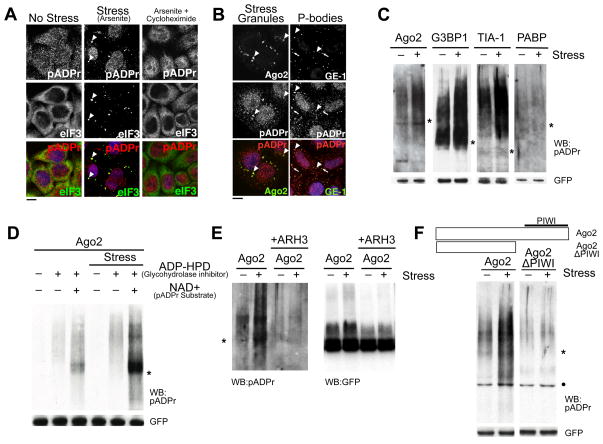
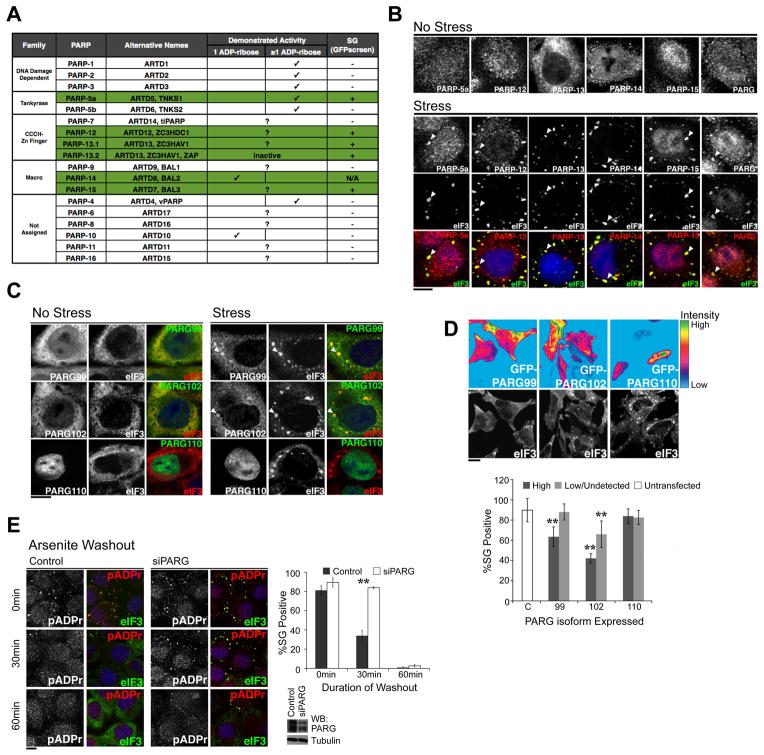
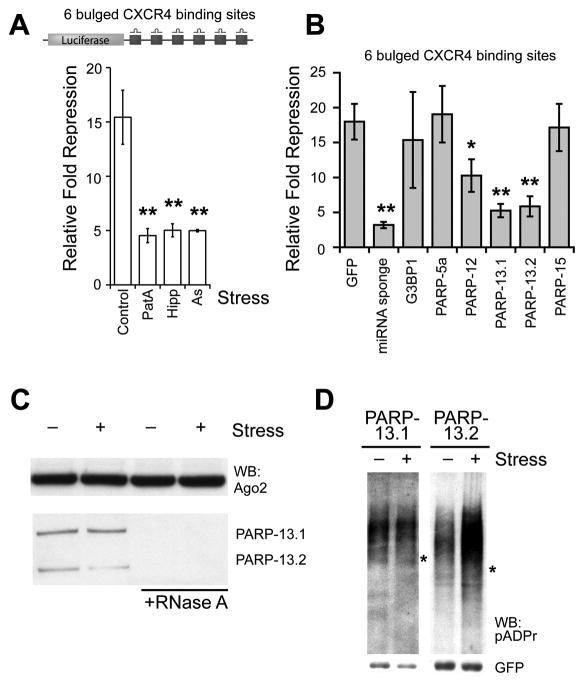
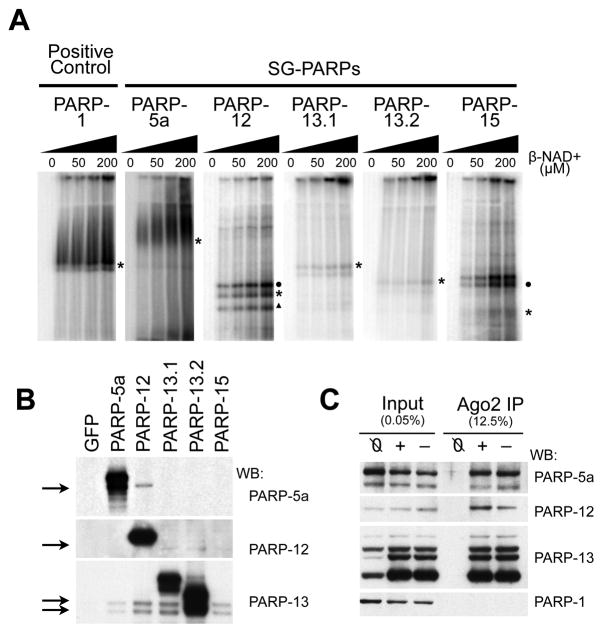
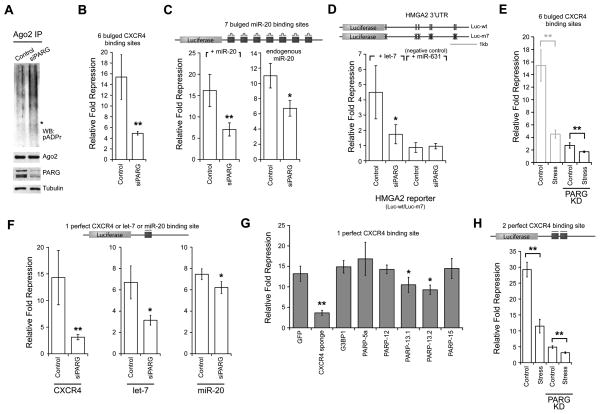
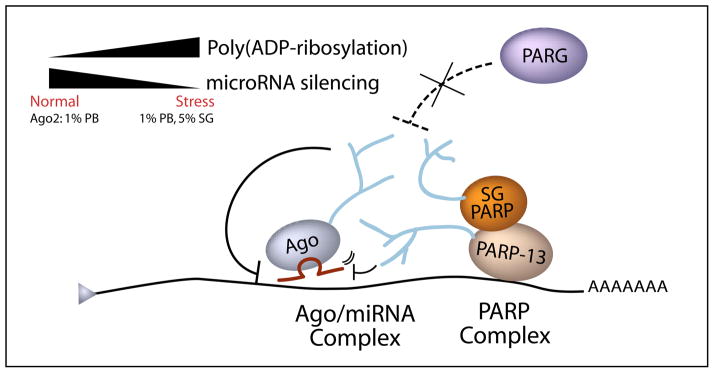
Poly(ADP-ribose) is a major regulatory macromolecule in the nucleus, where it regulates transcription, chromosome structure, and DNA damage repair. Functions in the interphase cytoplasm are less understood. Here, we identify a requirement for poly(ADP-ribose) in the assembly of cytoplasmic stress granules, which accumulate RNA-binding proteins that regulate the translation and stability of mRNAs upon stress. We show that poly(ADP-ribose), six specific poly(ADP-ribose) polymerases, and two poly(ADP-ribose) glycohydrolase isoforms are stress granule components. A subset of stress granule proteins, including microRNA-binding Argonaute family members Ago1-4, are modified by poly(ADP-ribose), and such modification increases upon stress, a condition when both microRNA-mediated translational repression and microRNA-directed mRNA cleavage are relieved. Similar relief of repression is also observed upon overexpression of specific poly(ADP-ribose) polymerases or, conversely, upon knockdown of glycohydrolase. We conclude that poly(ADP-ribose) is a key regulator of posttranscriptional gene expression in the cytoplasm.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Anderson P, Kedersha NL. Stress granules: the Tao of RNA triage. Trends Biochem Sci. 2008;33:141–150. - PubMed
-
- Bhattacharyya S, Habermacher R, Martine U, Closs E, Filipowicz W. Relief of microRNA-Mediated Translational Repression in Human Cells Subjected to Stress. Cell. 2006;125:1111–1124. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
