

Degrade, move, regroup: signaling control of splicing proteins
- PMID: 21596569
- PMCID: PMC3155649
- DOI: 10.1016/j.tibs.2011.04.003
Degrade, move, regroup: signaling control of splicing proteins
Abstract
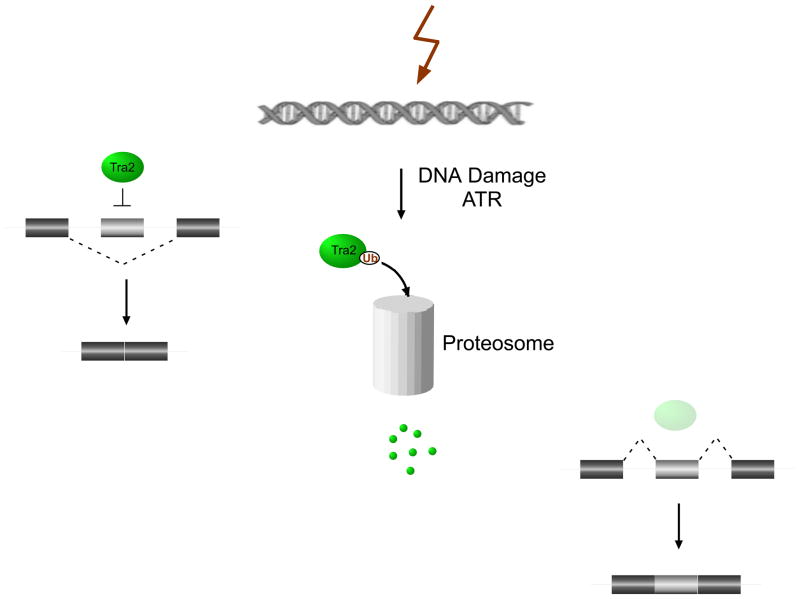
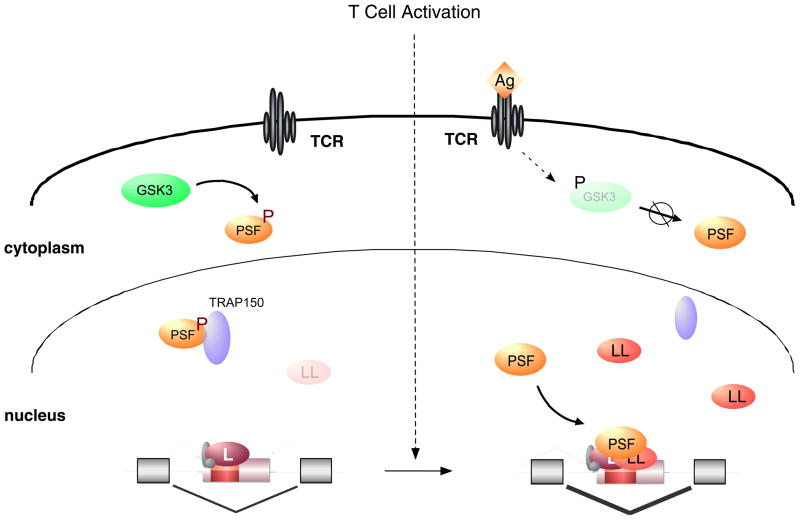
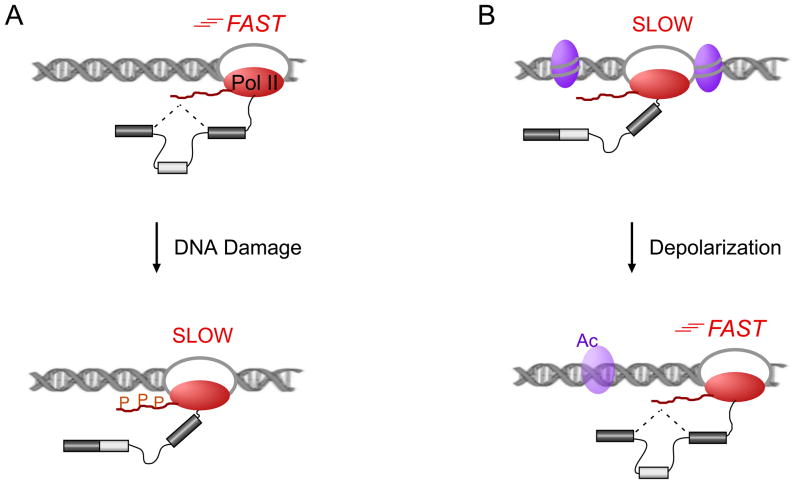
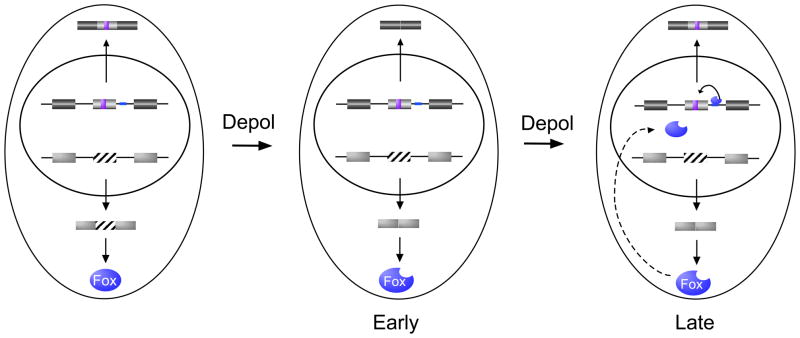
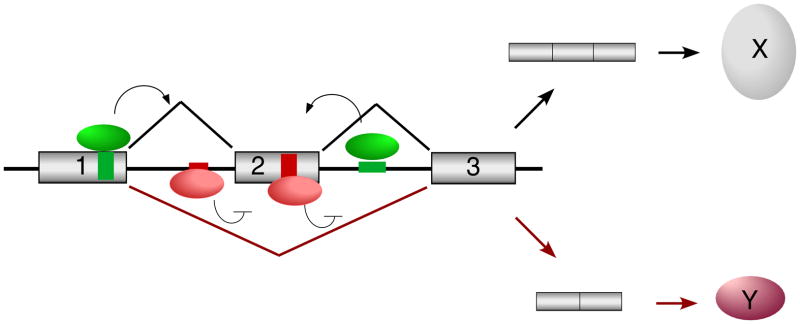
With recent advances in microarrays and sequencing it is now relatively straightforward to compare pre-mRNA splicing patterns in different cellular conditions on a genome-wide scale. Such studies have revealed extensive changes in cellular splicing programs in response to stimuli such as neuronal depolarization, DNA damage, immune signaling and cellular metabolic changes. However, for many years our understanding of the signaling pathways responsible for such splicing changes was greatly lacking. Excitingly, over the past few years this gap has begun to close. Recent studies now suggest notable trends in the mechanisms that link cellular stimuli to downstream alternative splicing events. These include regulated synthesis or degradation of splicing factors, differential protein-protein interactions, altered nuclear translocation and changes in transcription elongation.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Pan Q, et al. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nature genetics. 2008;40:1413–1415. - PubMed
-
- Lynch KW. Regulation of alternative splicing by signal transduction pathways. Adv Exp Med Biol. 2007;623:161–174. - PubMed
-
- Shin C, Manley JL. Cell signalling and the control of pre-mRNA splicing. Nature reviews. 2004;5:727–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
