

Phosphorylation of Mcm2 modulates Mcm2-7 activity and affects the cell's response to DNA damage
- PMID: 21596784
- PMCID: PMC3167627
- DOI: 10.1093/nar/gkr371
Phosphorylation of Mcm2 modulates Mcm2-7 activity and affects the cell's response to DNA damage
Abstract
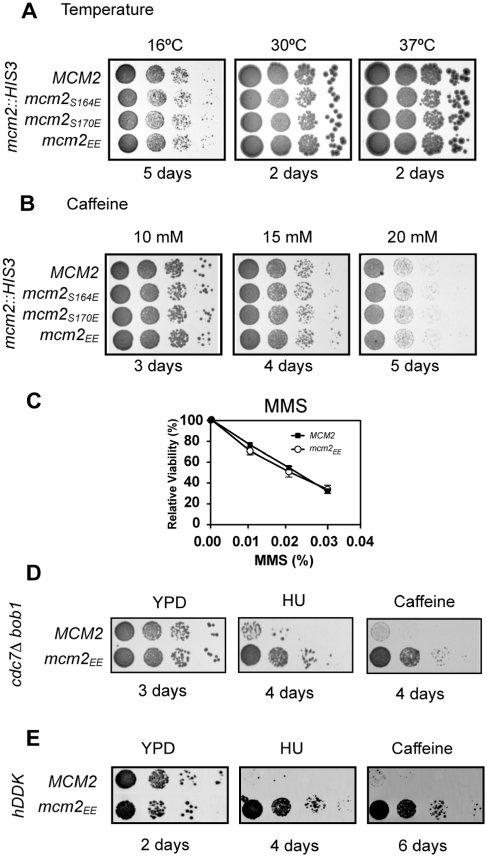
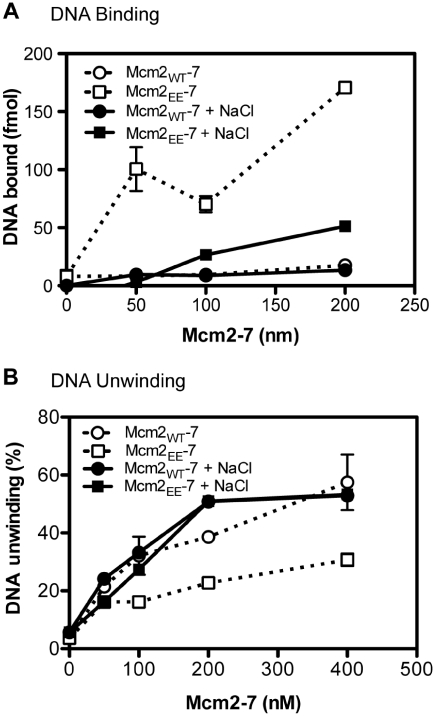
The S-phase kinase, DDK controls DNA replication through phosphorylation of the replicative helicase, Mcm2-7. We show that phosphorylation of Mcm2 at S164 and S170 is not essential for viability. However, the relevance of Mcm2 phosphorylation is demonstrated by the sensitivity of a strain containing alanine at these positions (mcm2(AA)) to methyl methanesulfonate (MMS) and caffeine. Consistent with a role for Mcm2 phosphorylation in response to DNA damage, the mcm2(AA) strain accumulates more RPA foci than wild type. An allele with the phosphomimetic mutations S164E and S170E (mcm2(EE)) suppresses the MMS and caffeine sensitivity caused by deficiencies in DDK function. In vitro, phosphorylation of Mcm2 or Mcm2(EE) reduces the helicase activity of Mcm2-7 while increasing DNA binding. The reduced helicase activity likely results from the increased DNA binding since relaxing DNA binding with salt restores helicase activity. The finding that the ATP site mutant mcm2(K549R) has higher DNA binding and less ATPase than mcm2(EE), but like mcm2(AA) results in drug sensitivity, supports a model whereby a specific range of Mcm2-7 activity is required in response to MMS and caffeine. We propose that phosphorylation of Mcm2 fine-tunes the activity of Mcm2-7, which in turn modulates DNA replication in response to DNA damage.
Figures






Similar articles
-
Mcm2 phosphorylation and the response to replicative stress.BMC Genet. 2012 May 7;13:36. doi: 10.1186/1471-2156-13-36. BMC Genet. 2012. PMID: 22564307 Free PMC article.
-
Dbf4 and Cdc7 proteins promote DNA replication through interactions with distinct Mcm2-7 protein subunits.J Biol Chem. 2013 May 24;288(21):14926-35. doi: 10.1074/jbc.M112.392910. Epub 2013 Apr 2. J Biol Chem. 2013. PMID: 23549044 Free PMC article.
-
Mec1 is one of multiple kinases that prime the Mcm2-7 helicase for phosphorylation by Cdc7.Mol Cell. 2010 Nov 12;40(3):353-63. doi: 10.1016/j.molcel.2010.10.017. Mol Cell. 2010. PMID: 21070963 Free PMC article.
-
Getting a grip on licensing: mechanism of stable Mcm2-7 loading onto replication origins.Mol Cell. 2006 Jan 20;21(2):143-4. doi: 10.1016/j.molcel.2006.01.003. Mol Cell. 2006. PMID: 16427002 Review.
-
The Eukaryotic CMG Helicase at the Replication Fork: Emerging Architecture Reveals an Unexpected Mechanism.Bioessays. 2018 Mar;40(3):10.1002/bies.201700208. doi: 10.1002/bies.201700208. Epub 2018 Feb 6. Bioessays. 2018. PMID: 29405332 Free PMC article. Review.
Cited by
-
DNA replication: Mechanisms and therapeutic interventions for diseases.MedComm (2020). 2023 Feb 5;4(1):e210. doi: 10.1002/mco2.210. eCollection 2023 Feb. MedComm (2020). 2023. PMID: 36776764 Free PMC article. Review.
-
Insights into the Initiation of Eukaryotic DNA Replication.Nucleus. 2015;6(6):449-54. doi: 10.1080/19491034.2015.1115938. Epub 2015 Dec 28. Nucleus. 2015. PMID: 26710261 Free PMC article.
-
The Minichromosome Maintenance Complex Component 2 (MjMCM2) of Meloidogyne javanica is a potential effector regulating the cell cycle in nematode-induced galls.Sci Rep. 2022 Jun 2;12(1):9196. doi: 10.1038/s41598-022-13020-8. Sci Rep. 2022. PMID: 35654810 Free PMC article.
-
Mcm2 phosphorylation and the response to replicative stress.BMC Genet. 2012 May 7;13:36. doi: 10.1186/1471-2156-13-36. BMC Genet. 2012. PMID: 22564307 Free PMC article.
-
Conserved mechanism for coordinating replication fork helicase assembly with phosphorylation of the helicase.Proc Natl Acad Sci U S A. 2015 Sep 8;112(36):11223-8. doi: 10.1073/pnas.1509608112. Epub 2015 Aug 24. Proc Natl Acad Sci U S A. 2015. PMID: 26305950 Free PMC article.
References
-
- Davey MJ, Indiani C, O'Donnell M. Reconstitution of the Mcm2-7p heterohexamer, subunit arrangement, and ATP site architecture. J. Biol. Chem. 2003;278:4491–4499. - PubMed
-
- Schwacha A, Bell SP. Interactions between two catalytically distinct MCM subgroups are essential for coordinated ATP hydrolysis and DNA replication. Mol. Cell. 2001;8:1093–1104. - PubMed
-
- Bochman ML, Schwacha A. The Mcm2-7 Complex has in vitro helicase activity. Mol. Cell. 2008;31:287–293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
