

Canonical hedgehog signaling augments tumor angiogenesis by induction of VEGF-A in stromal perivascular cells
- PMID: 21597001
- PMCID: PMC3111273
- DOI: 10.1073/pnas.1017945108
Canonical hedgehog signaling augments tumor angiogenesis by induction of VEGF-A in stromal perivascular cells
Abstract
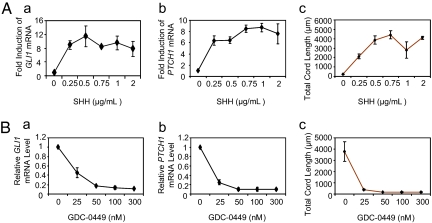
Hedgehog (Hh) signaling is critical to the patterning and development of a variety of organ systems, and both ligand-dependent and ligand-independent Hh pathway activation are known to promote tumorigenesis. Recent studies have shown that in tumors promoted by Hh ligands, activation occurs within the stromal microenvironment. Testing whether ligand-driven Hh signaling promotes tumor angiogenesis, we found that Hh antagonism reduced the vascular density of Hh-producing LS180 and SW480 xenografts. In addition, ectopic expression of sonic hedgehog in low-Hh-expressing DLD-1 xenografts increased tumor vascular density, augmented angiogenesis, and was associated with canonical Hh signaling within perivascular tumor stromal cells. To better understand the molecular mechanisms underlying Hh-mediated tumor angiogenesis, we established an Hh-sensitive angiogenesis coculture assay and found that fibroblast cell lines derived from a variety of human tissues were Hh responsive and promoted angiogenesis in vitro through a secreted paracrine signal(s). Affymetrix array analyses of cultured fibroblasts identified VEGF-A, hepatocyte growth factor, and PDGF-C as candidate secreted proangiogenic factors induced by Hh stimulation. Expression studies of xenografts and angiogenesis assays using combinations of Hh and VEGF-A inhibitors showed that it is primarily Hh-induced VEGF-A that promotes angiogenesis in vitro and augments tumor-derived VEGF to promote angiogenesis in vivo.
Conflict of interest statement
Conflict of interest statement: All authors are employees of Genentech, Inc., a wholly owned subsidiary of F. Hoffmann-La Roche Ltd., and may own equity in Roche.
Figures



References
-
- Scales SJ, de Sauvage FJ. Mechanisms of Hedgehog pathway activation in cancer and implications for therapy. Trends Pharmacol Sci. 2009;30:303–312. - PubMed
-
- Yauch RL, et al. A paracrine requirement for hedgehog signalling in cancer. Nature. 2008;455:406–410. - PubMed
-
- Byrd N, Grabel L. Hedgehog signaling in murine vasculogenesis and angiogenesis. Trends Cardiovasc Med. 2004;14:308–313. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
