

Vorinostat modulates cell cycle regulatory proteins in glioma cells and human glioma slice cultures
- PMID: 21598070
- PMCID: PMC3794467
- DOI: 10.1007/s11060-011-0604-7
Vorinostat modulates cell cycle regulatory proteins in glioma cells and human glioma slice cultures
Abstract
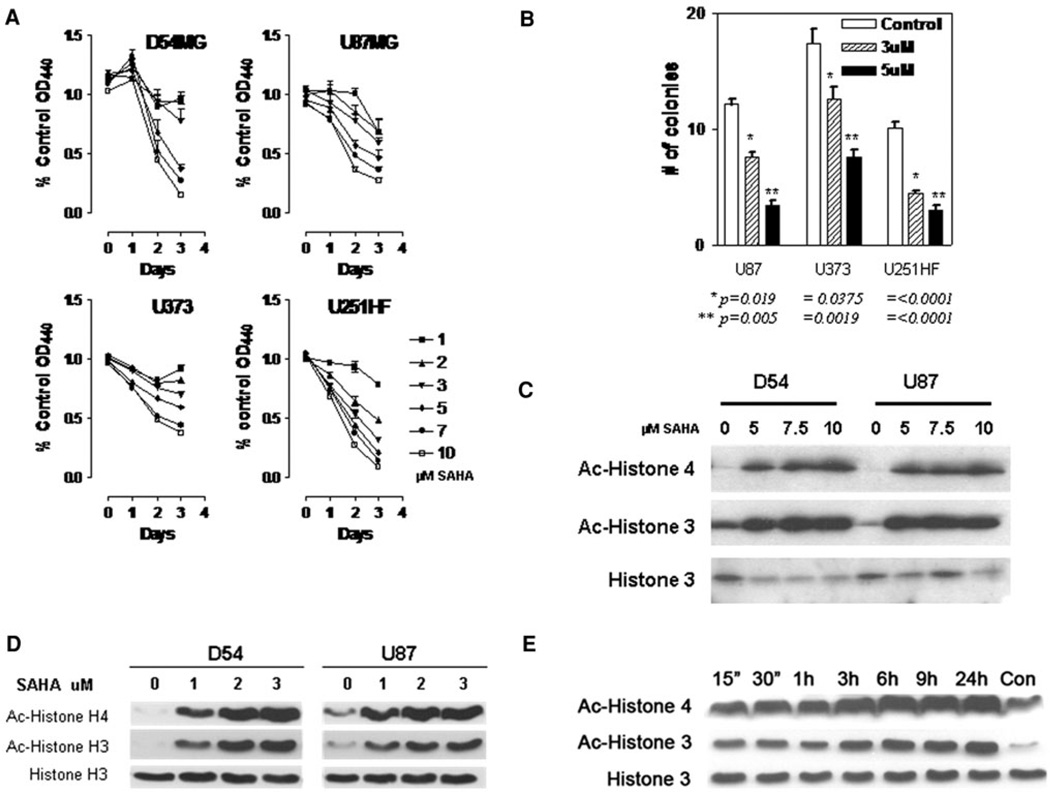
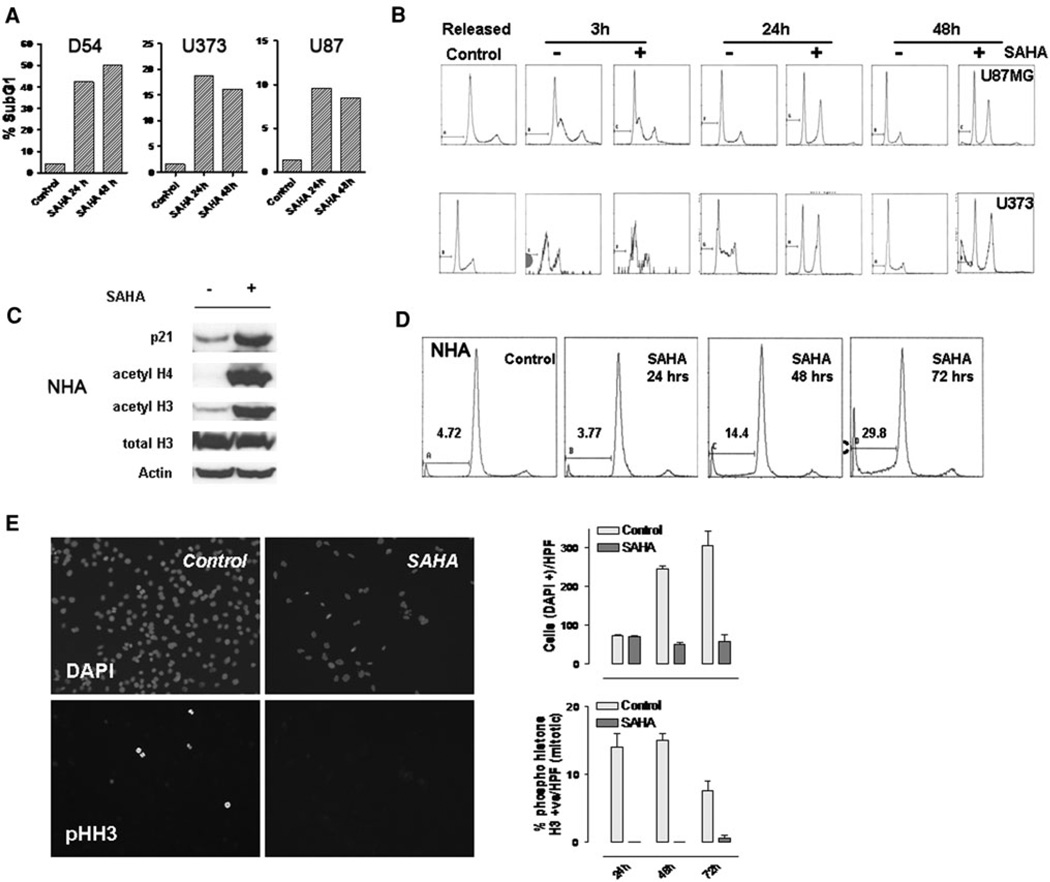
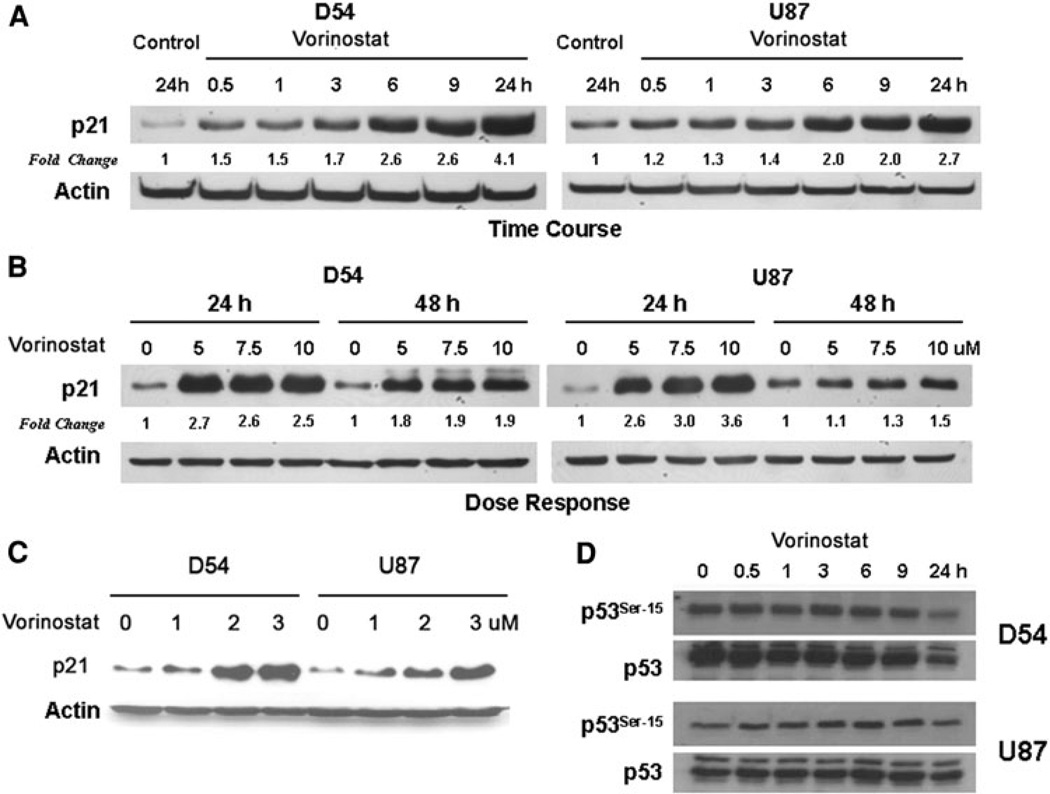
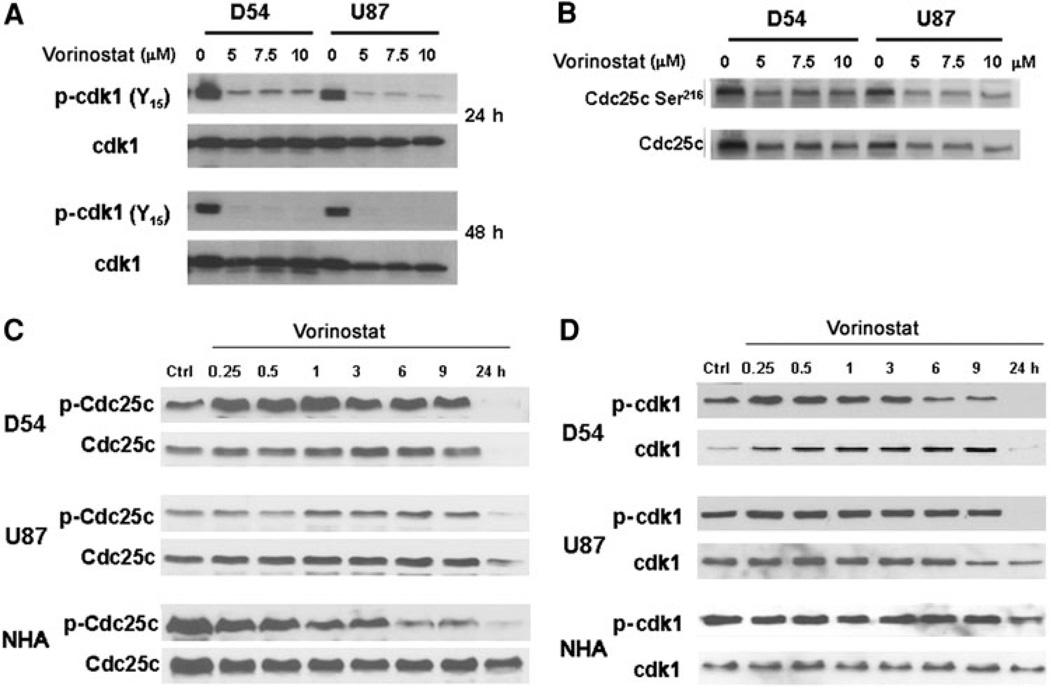
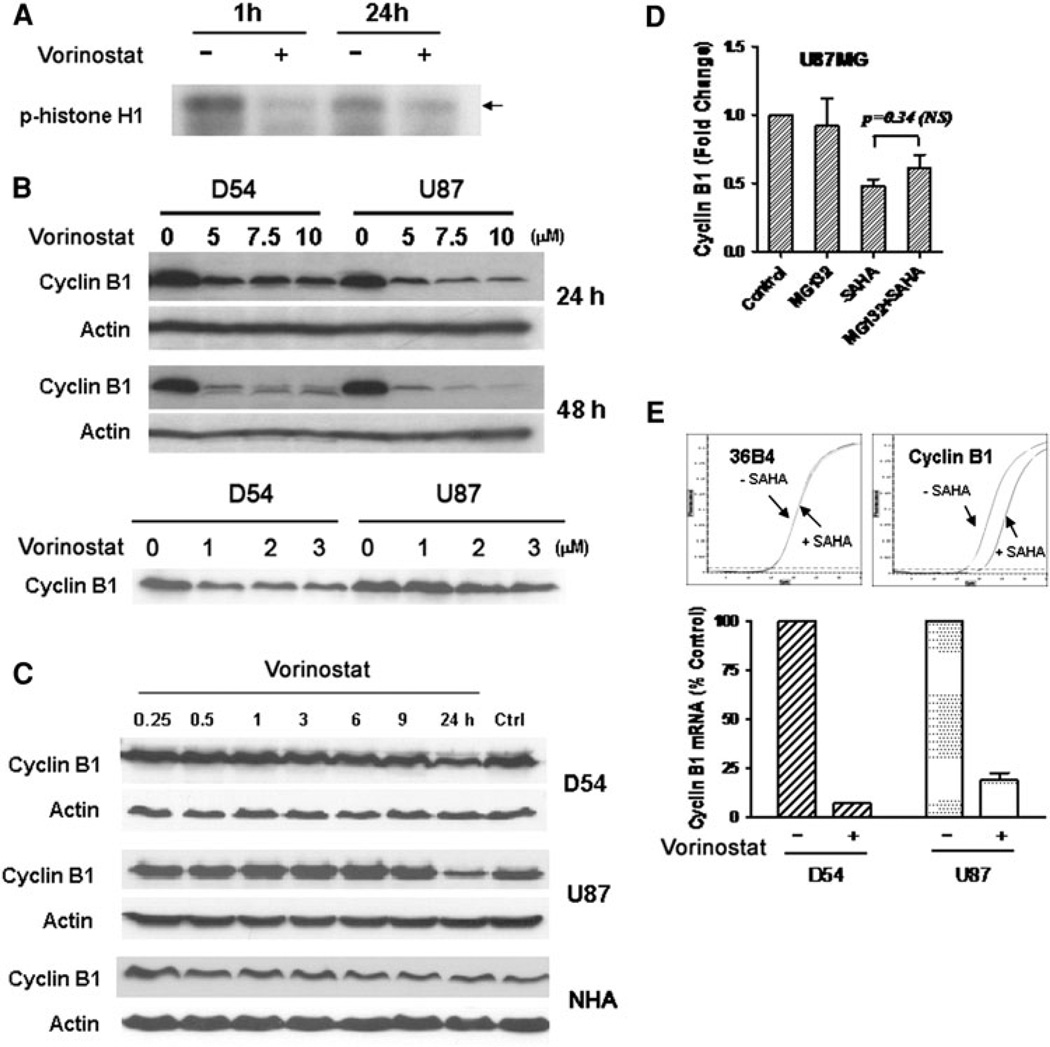
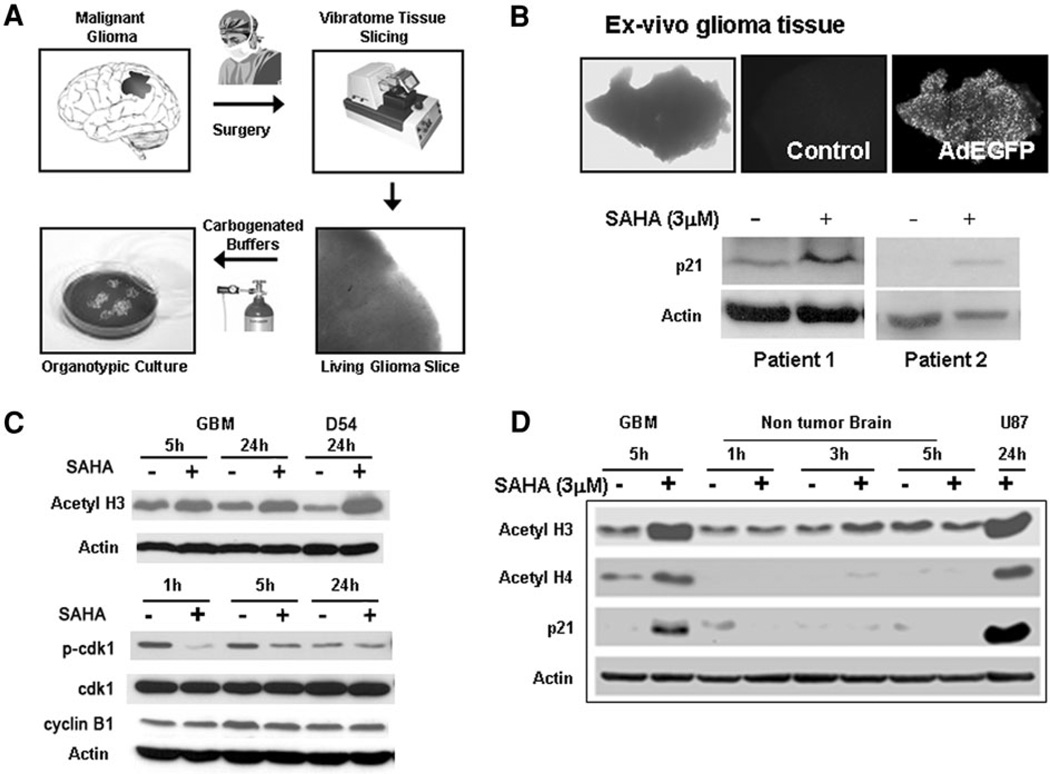
Chromatin modification through histone deacetylase inhibition has shown evidence of activity against malignancies. The mechanism of action of such agents are pleiotropic and potentially tumor specific. In this study, we studied the mechanisms of vorinostat-induced cellular effects in gliomas. The effects of vorinostat on proliferation, induction of apoptosis and cell cycle effects were studied in vitro (D54, U87 and U373 glioma cell lines). To gain additional insights into its effects on human gliomas, vorinostat-induced changes were examined ex vivo using a novel organotypic human glioma slice model. Vorinostat treatment resulted in increased p21 levels in all glioma cells tested in a p53 independent manner. In addition, cyclin B1 levels were transcriptionally downregulated and resulted in reduced kinase activity of the cyclin B1/cdk1 complex causing a G2 arrest. These effects were associated with a dose- and time-dependent inhibition of cellular proliferation and anchorage-independent growth in association with hyperacetylation of core histones and induction of apoptosis. Of particular significance, we demonstrate histone hyperacetylation and increased p21 levels in freshly resected human glioma specimens maintained as organotypic slice cultures and exposed to vorinostat similar to cell lines suggesting that human glioma can be targeted by this agent. Our data suggest that the effects of vorinostat are associated with modulation of cell cycle related proteins and activation of a G2 checkpoint along with induction of apoptosis. These effects are mediated by both transcriptional and post-translational mechanisms which provide potential options that can be exploited to develop new therapeutic approaches against gliomas.
Conflict of interest statement
Figures
References
-
- Holland EC. Gliomagenesis: genetic alterations and mouse models. Nat Rev. 2001;2:120–129. - PubMed
-
- Penas-Prado M, Gilbert MR. Molecularly targeted therapies for malignant gliomas: advances and challenges. Expert Rev Anticancer Ther. 2007;7:641–661. - PubMed
-
- Baylin SB. DNA methylation and gene silencing in cancer. Nat Clin Pract. 2005;2(Suppl 1):S4–S11. - PubMed
-
- Amatya VJ, Naumann U, Weller M, Ohgaki H. TP53 promoter methylation in human gliomas. Acta Neuropathol. 2005;110:178–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
