

Timing the tides: genetic control of diurnal and lunar emergence times is correlated in the marine midge Clunio marinus
- PMID: 21599938
- PMCID: PMC3124415
- DOI: 10.1186/1471-2156-12-49
Timing the tides: genetic control of diurnal and lunar emergence times is correlated in the marine midge Clunio marinus
Abstract
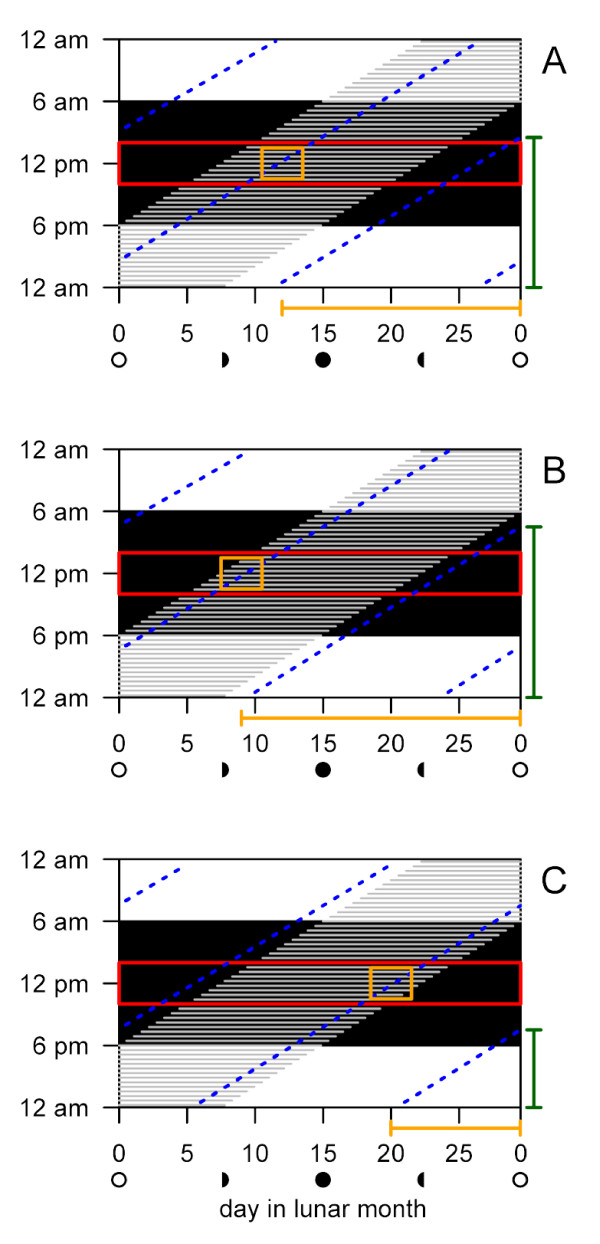
Background: The intertidal zone of seacoasts, being affected by the superimposed tidal, diurnal and lunar cycles, is temporally the most complex environment on earth. Many marine organisms exhibit lunar rhythms in reproductive behaviour and some show experimental evidence of endogenous control by a circalunar clock, the molecular and genetic basis of which is unexplored. We examined the genetic control of lunar and diurnal rhythmicity in the marine midge Clunio marinus (Chironomidae, Diptera), a species for which the correct timing of adult emergence is critical in natural populations.
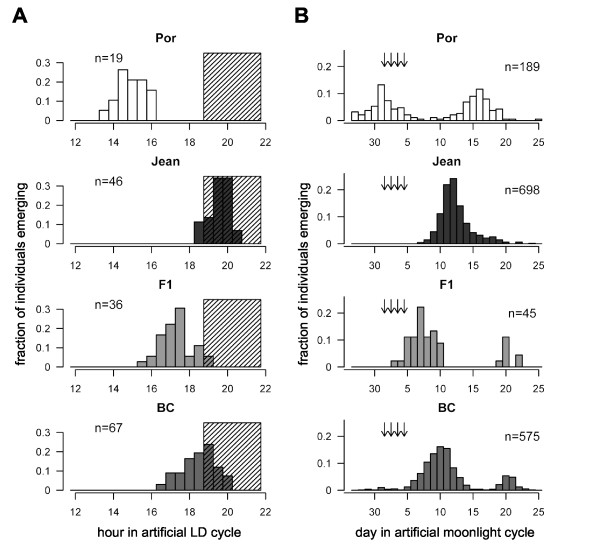
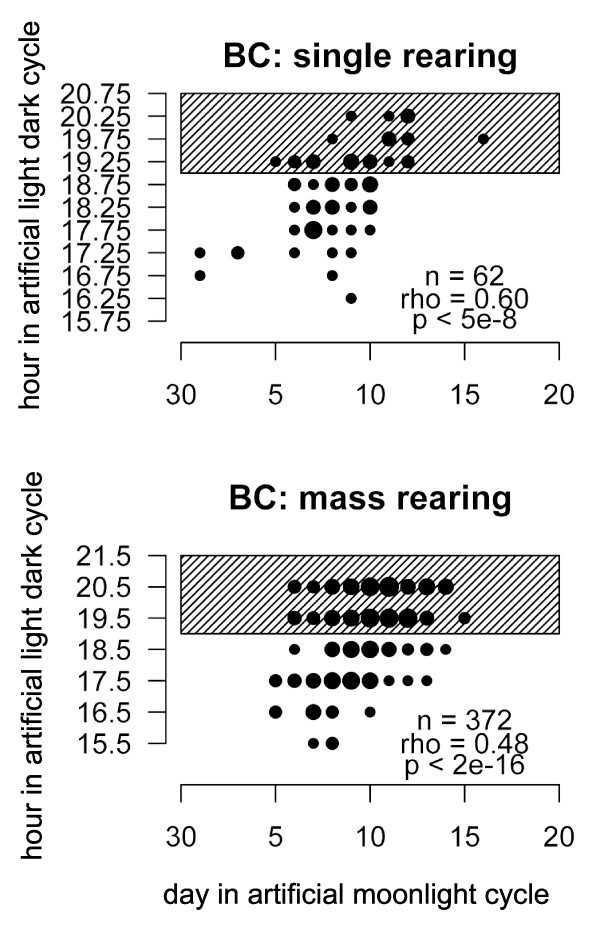
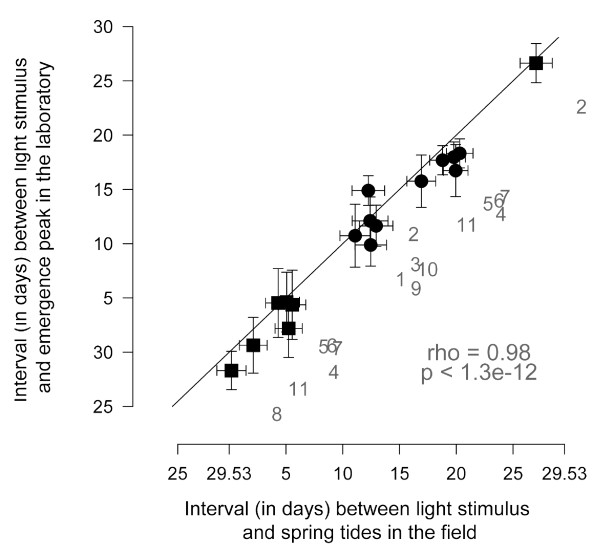
Results: We crossed two strains of Clunio marinus that differ in the timing of the diurnal and lunar rhythms of emergence. The phenotype distribution of the segregating backcross progeny indicates polygenic control of the lunar emergence rhythm. Diurnal timing of emergence is also under genetic control, and is influenced by two unlinked genes with major effects. Furthermore, the lunar and diurnal timing of emergence is correlated in the backcross generation. We show that both the lunar emergence time and its correlation to the diurnal emergence time are adaptive for the species in its natural environment.
Conclusions: The correlation implies that the unlinked genes affecting lunar timing and the two unlinked genes affecting diurnal timing could be the same, providing an unexpectedly close interaction of the two clocks. Alternatively, the genes could be genetically linked in a two-by-two fashion, suggesting that evolution has shaped the genetic architecture to stabilize adaptive combinations of lunar and diurnal emergence times by tightening linkage. Our results, the first on genetic control of lunar rhythms, offer a new perspective to explore their molecular clockwork.
Figures
References
-
- Franke HD. The role of light and endogenous factors in the timing of the reproductive-cycle of Typosyllis prolifera and some other polychaetes. American Zoologist. 1986;26(2):433–445.
-
- Hauenschild C. Lunar Periodicity. Cold Spring Harbor Symposia on Quantitative Biology. 1960;25:491–497. - PubMed
-
- Naylor E. Marine animal behaviour in relation to lunar phase. Earth Moon and Planets. 2001;85-6:291–302.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
