

Interplay between flavodiiron proteins and photorespiration in Synechocystis sp. PCC 6803
- PMID: 21602273
- PMCID: PMC3129182
- DOI: 10.1074/jbc.M111.223289
Interplay between flavodiiron proteins and photorespiration in Synechocystis sp. PCC 6803
Abstract
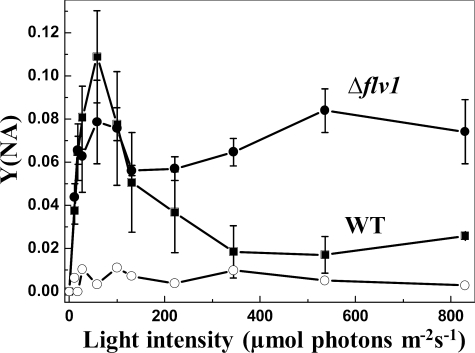
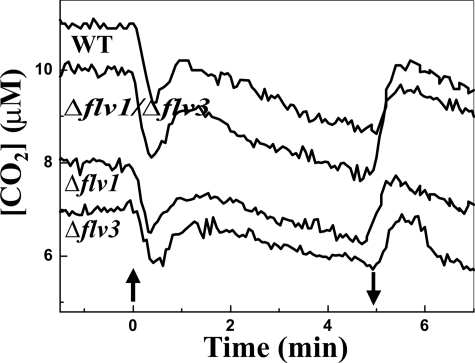
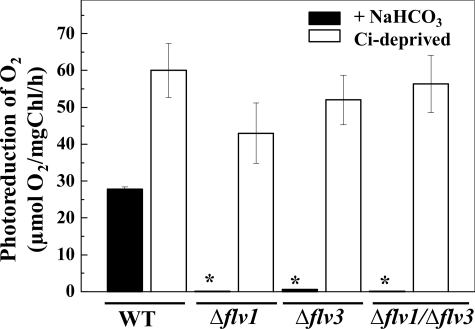
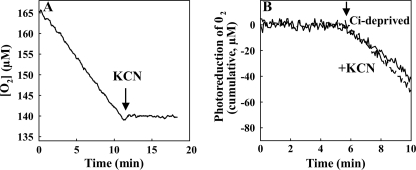
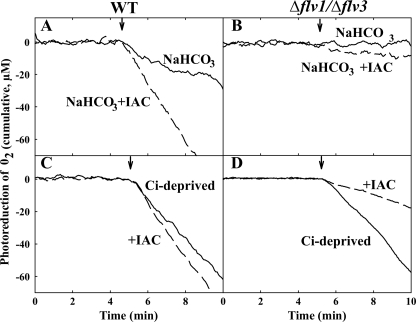
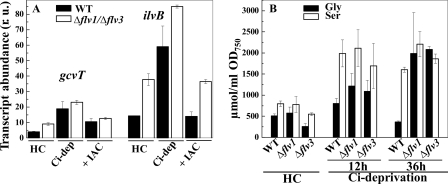
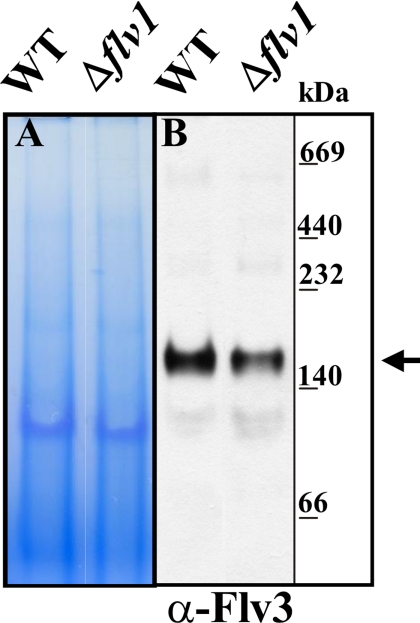
Flavodiiron (Flv) proteins are involved in detoxification of O(2) and NO in anaerobic bacteria and archaea. Cyanobacterial Flv proteins, on the contrary, function in oxygenic environment and possess an extra NAD(P)H:flavin oxidoreductase module. Synechocystis sp. PCC 6803 has four genes (sll1521, sll0219, sll0550, and sll0217) encoding Flv proteins (Flv1, Flv2, Flv3, and Flv4). Previous in vitro studies with recombinant Flv3 protein from Synechocystis provided evidence that it functions as a NAD(P)H:oxygen oxidoreductase, and subsequent in vivo studies with Synechocystis confirmed the role of Flv1 and Flv3 proteins in the Mehler reaction (photoreduction of O(2) to H(2)O). Interestingly, homologous proteins to Flv1 and Flv3 can be found also in green algae, mosses, and Selaginella. Here, we addressed the function of Flv1 and Flv3 in Synechocystis using the Δflv1, Δflv3, and Δflv1/Δflv3 mutants and applying inorganic carbon (C(i))-deprivation conditions. We propose that only the Flv1/Flv3 heterodimer form is functional in the Mehler reaction in vivo. (18)O(2) labeling was used to discriminate between O(2) evolution in photosynthetic water splitting and O(2) consumption. In wild type, ∼20% of electrons originated from water was targeted to O(2) under air level CO(2) conditions but increased up to 60% in severe limitation of C(i). Gas exchange experiments with Δflv1, Δflv3, and Δflv1/Δflv3 mutants demonstrated that a considerable amount of electrons in these mutants is directed to photorespiration under C(i) deprivation. This assumption is in line with increased transcript abundance of photorespiratory genes and accumulation of photorespiratory intermediates in the WT and to a higher extent in mutant cells under C(i) deprivation.
Figures
References
-
- Vicente J. B., Justino M. C., Gonçalves V. L., Saraiva L. M., Teixeira M. (2008) Methods Enzymol. 437, 21–45 - PubMed
-
- Vicente J. B., Testa F., Mastronicola D., Forte E., Sarti P., Teixeira M., Giuffrè A. (2009) Arch. Biochem. Biophys. 488, 9–13 - PubMed
-
- Di Matteo A., Scandurra F. M., Testa F., Forte E., Sarti P., Brunori M., Giuffrè A. (2008) J. Biol. Chem. 283, 4061–4068 - PubMed
-
- Frazão C., Silva G., Gomes C. M., Matias P., Coelho R., Sieker L., Macedo S., Liu M. Y., Oliveira S., Teixeira M., Xavier A. V., Rodrigues-Pousada C., Carrondo M. A., Le Gall J. (2000) Nat. Struct. Biol. 7, 1041–1045 - PubMed
-
- Vicente J. B., Gomes C. M., Wasserfallen A., Teixeira M. (2002) Biochem. Biophys. Res. Commun. 294, 82–87 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
