

FadD19 of Rhodococcus rhodochrous DSM43269, a steroid-coenzyme A ligase essential for degradation of C-24 branched sterol side chains
- PMID: 21602385
- PMCID: PMC3127730
- DOI: 10.1128/AEM.00380-11
FadD19 of Rhodococcus rhodochrous DSM43269, a steroid-coenzyme A ligase essential for degradation of C-24 branched sterol side chains
Abstract
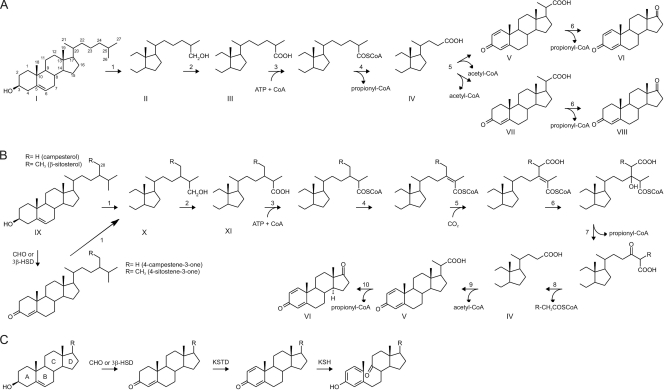
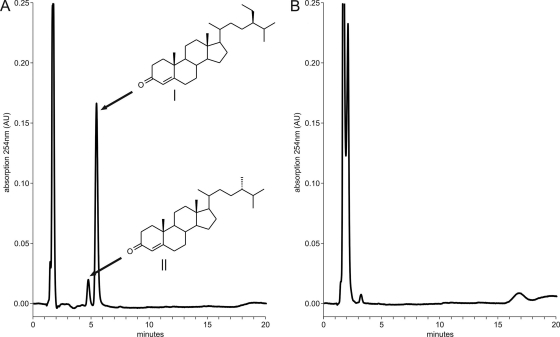
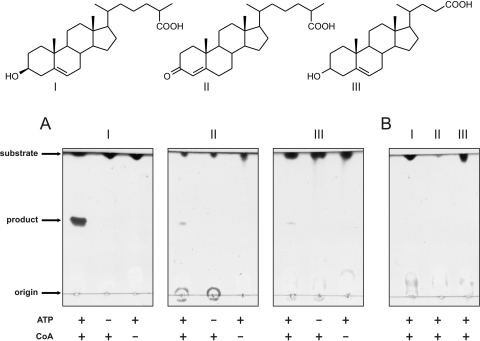
The actinobacterial cholesterol catabolic gene cluster contains a subset of genes that encode β-oxidation enzymes with a putative role in sterol side chain degradation. We investigated the physiological roles of several genes, i.e., fadD17, fadD19, fadE26, fadE27, and ro04690DSM43269, by gene inactivation studies in mutant strain RG32 of Rhodococcus rhodochrous DSM43269. Mutant strain RG32 is devoid of 3-ketosteroid 9α-hydroxylase (KSH) activity and was constructed following the identification, cloning, and sequential inactivation of five kshA gene homologs in strain DSM43269. We show that mutant strain RG32 is fully blocked in steroid ring degradation but capable of selective sterol side chain degradation. Except for RG32ΔfadD19, none of the mutants constructed in RG32 revealed an aberrant phenotype on sterol side chain degradation compared to parent strain RG32. Deletion of fadD19 in strain RG32 completely blocked side chain degradation of C-24 branched sterols but interestingly not that of cholesterol. The additional inactivation of fadD17 in mutant RG32ΔfadD19 also did not affect cholesterol side chain degradation. Heterologously expressed FadD19DSM43269 nevertheless was active toward steroid-C26-oic acid substrates. Our data identified FadD19 as a steroid-coenzyme A (CoA) ligase with an essential in vivo role in the degradation of the side chains of C-24 branched-chain sterols. This paper reports the identification and characterization of a CoA ligase with an in vivo role in sterol side chain degradation. The high similarity (67%) between the FadD19(DSM43269) and FadD19H37Rv enzymes further suggests that FadD19H37Rv has an in vivo role in sterol metabolism of Mycobacterium tuberculosis H37Rv.
Figures

References
-
- Arima K., Nagasawa M., Bae M., Tamura G. 1969. Microbial transformation of sterols. Part I. Decomposition of cholesterol by microorganisms. Agric. Biol. Chem. 33:1636–1643
-
- Arima K., Nakamatsu T., Beppu T. 1978. Microbial production of 3-oxobisnorchola-1,4-dien-22-oic acid. Agric. Biol. Chem. 42:411–416
-
- Arora P., Vats A., Saxena P., Mohanty D., Gokhale R. S. 2005. Promiscuous fatty acyl CoA ligases produce acyl-CoA and acyl-SNAC precursors for polyketide biosynthesis. J. Am. Chem. Soc. 127:9388–9389 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
