

Pathogenesis of Candida albicans infections in the alternative chorio-allantoic membrane chicken embryo model resembles systemic murine infections
- PMID: 21603634
- PMCID: PMC3094387
- DOI: 10.1371/journal.pone.0019741
Pathogenesis of Candida albicans infections in the alternative chorio-allantoic membrane chicken embryo model resembles systemic murine infections
Erratum in
- PLoS One. 2011;6(6). doi:10.1371/annotation/1912450a-b5a7-4dbe-bc2a-cd93579fb0dd
Abstract
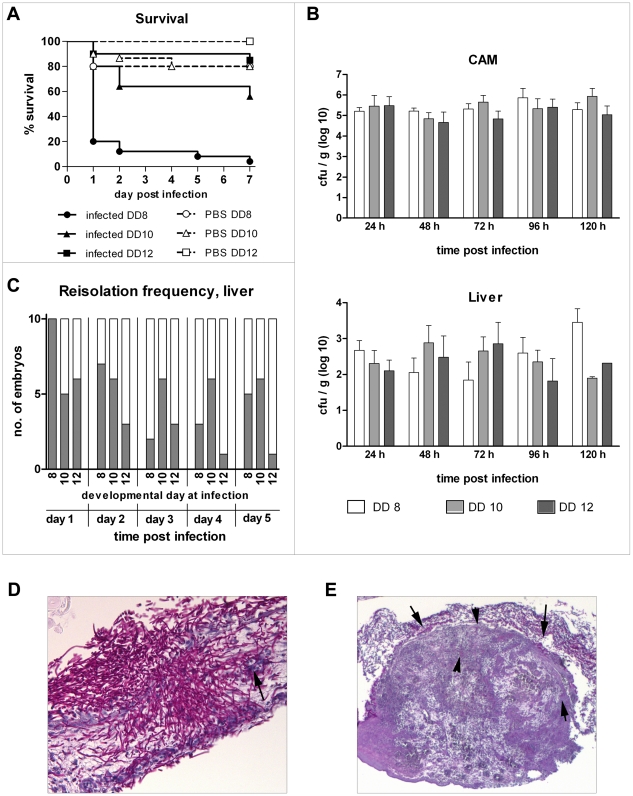
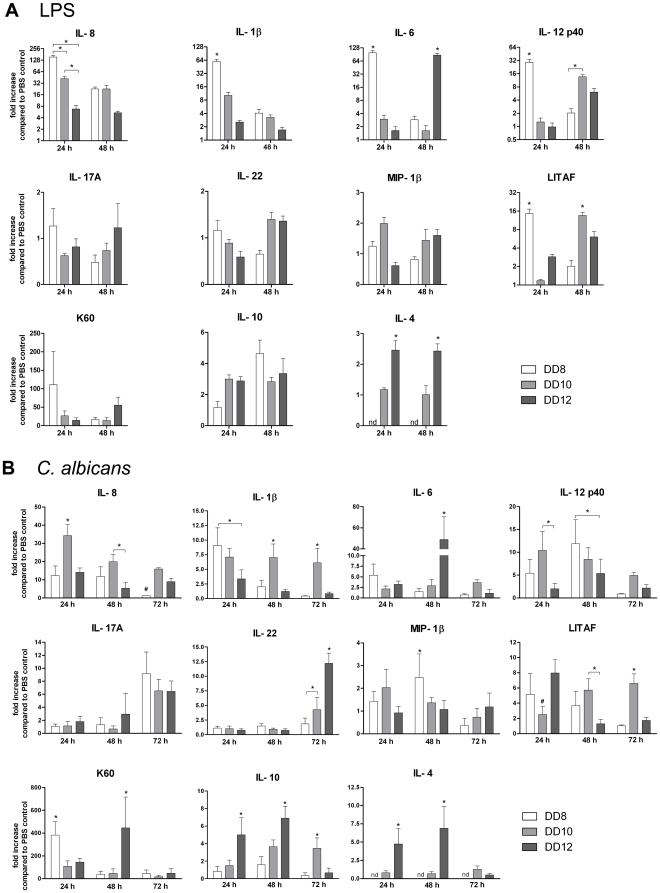
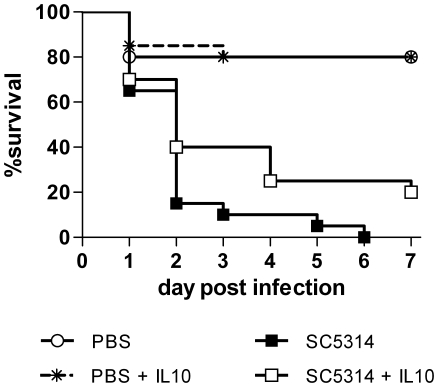
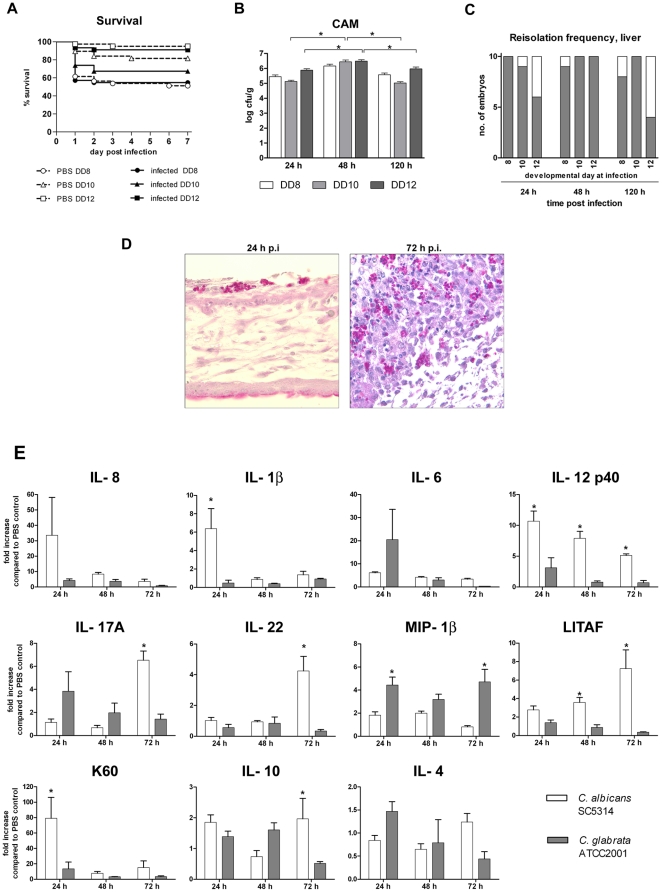
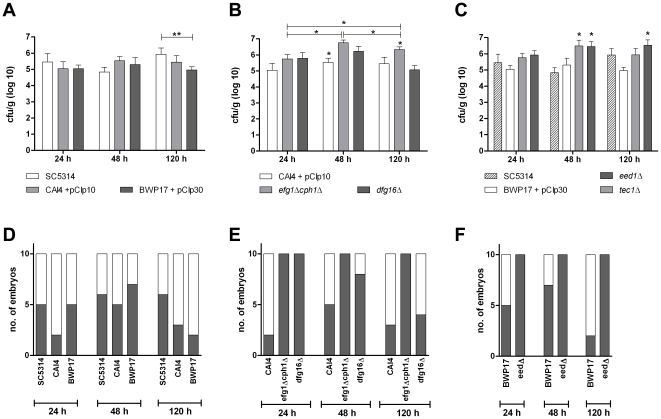
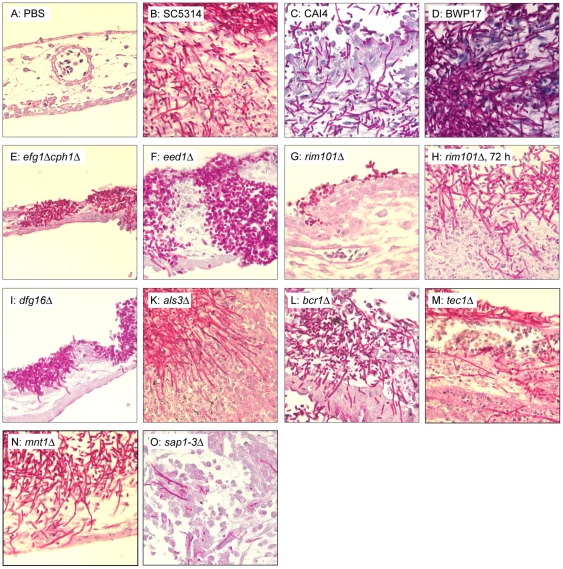
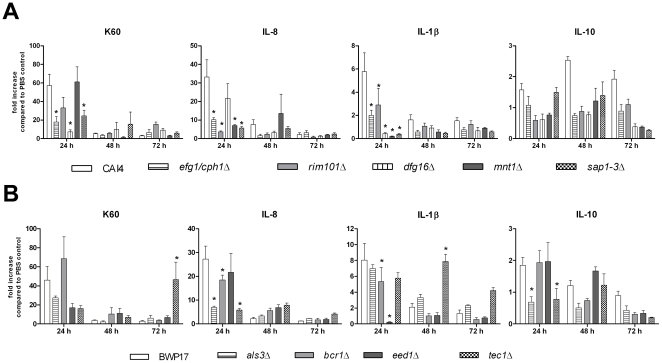
Alternative models of microbial infections are increasingly used to screen virulence determinants of pathogens. In this study, we investigated the pathogenesis of Candida albicans and C. glabrata infections in chicken embryos infected via the chorio-allantoic membrane (CAM) and analyzed the virulence of deletion mutants. The developing immune system of the host significantly influenced susceptibility: With increasing age, embryos became more resistant and mounted a more balanced immune response, characterized by lower induction of proinflammatory cytokines and increased transcription of regulatory cytokines, suggesting that immunopathology contributes to pathogenesis. While many aspects of the chicken embryo response resembled murine infections, we also observed significant differences: In contrast to systemic infections in mice, IL-10 had a beneficial effect in chicken embryos. IL-22 and IL-17A were only upregulated after the peak mortality in the chicken embryo model occurred; thus, the role of the Th17 response in this model remains unclear. Abscess formation occurs frequently in murine models, whereas the avian response was dominated by granuloma formation. Pathogenicity of the majority of 15 tested C. albicans deletion strains was comparable to the virulence in mouse models and reduced virulence was associated with significantly lower transcription of proinflammatory cytokines. However, fungal burden did not correlate with virulence and for few mutants like bcr1Δ and tec1Δ different outcomes in survival compared to murine infections were observed. C. albicans strains locked in the yeast stage disseminated significantly more often from the CAM into the embryo, supporting the hypothesis that the yeast morphology is responsible for dissemination in systemic infections. These data suggest that the pathogenesis of C. albicans infections in the chicken embryo model resembles systemic murine infections but also differs in some aspects. Despite its limitations, it presents a useful alternative tool to pre-screen C. albicans strains to select strains for subsequent testing in murine models.
Conflict of interest statement
Figures
References
-
- Odds FC. Candida and Candidosis. London: Baillière Tindall; 1988.
-
- Pappas PG, Rex JH, Lee J, Hamill RJ, Larsen RA, et al. A prospective observational study of candidemia: epidemiology, therapy, and influences on mortality in hospitalized adult and pediatric patients. Clin Infect Dis. 2003;37:634–643. - PubMed
-
- de Repentigny L. Animal models in the analysis of Candida host-pathogen interactions. Curr Opin Microbiol. 2004;7:324–329. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
