

Farnesyl pyrophosphate regulates adipocyte functions as an endogenous PPARγ agonist
- PMID: 21605082
- PMCID: PMC3253524
- DOI: 10.1042/BJ20101939
Farnesyl pyrophosphate regulates adipocyte functions as an endogenous PPARγ agonist
Abstract
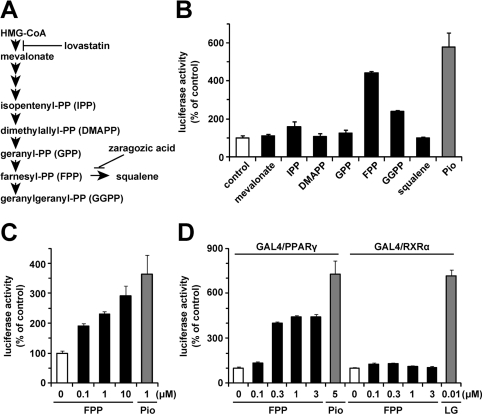
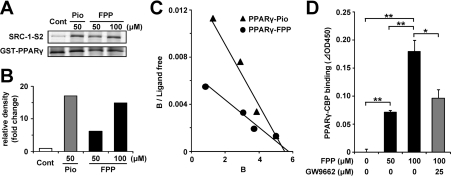
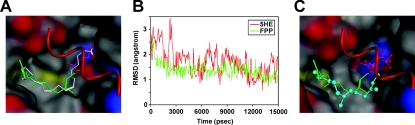
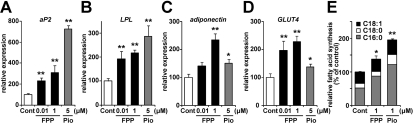
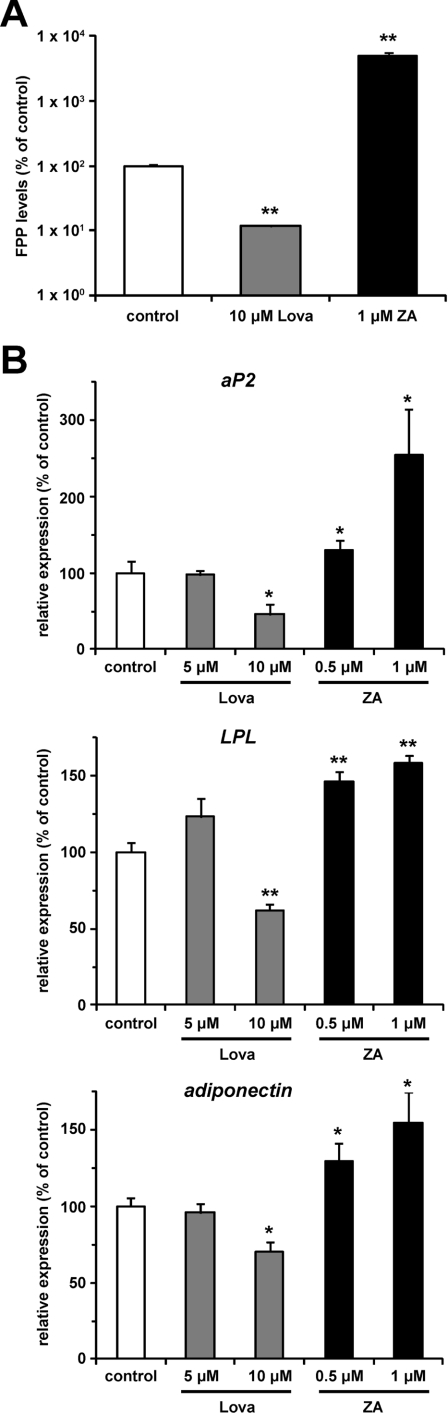
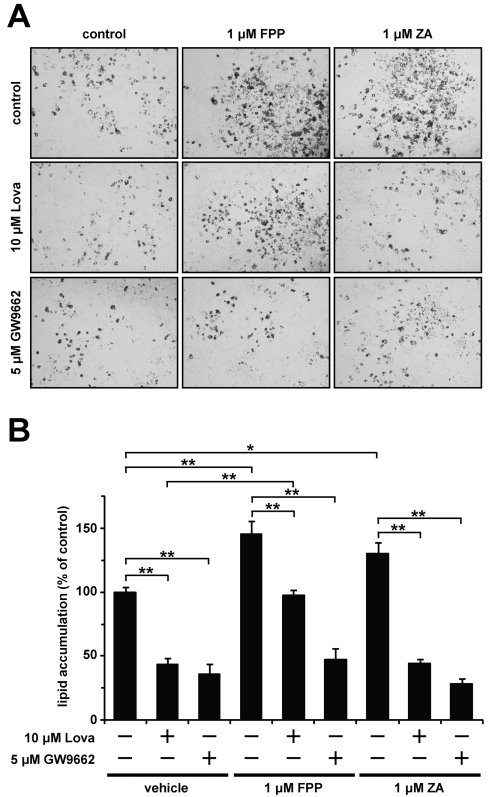
The cholesterol biosynthetic pathway produces not only sterols but also non-sterol mevalonate metabolites involved in isoprenoid synthesis. Mevalonate metabolites affect transcriptional and post-transcriptional events that in turn affect various biological processes including energy metabolism. In the present study, we examine whether mevalonate metabolites activate PPARγ (peroxisome-proliferator-activated receptor γ), a ligand-dependent transcription factor playing a central role in adipocyte differentiation. In the luciferase reporter assay using both GAL4 chimaera and full-length PPARγ systems, a mevalonate metabolite, FPP (farnesyl pyrophosphate), which is the precursor of almost all isoprenoids and is positioned at branch points leading to the synthesis of other longer-chain isoprenoids, activated PPARγ in a dose-dependent manner. FPP induced the in vitro binding of a co-activator, SRC-1 (steroid receptor co-activator-1), to GST (glutathione transferase)-PPARγ. Direct binding of FPP to PPARγ was also indicated by docking simulation studies. Moreover, the addition of FPP up-regulated the mRNA expression levels of PPARγ target genes during adipocyte differentiation induction. In the presence of lovastatin, an HMG-CoA (3-hydroxy-3-methylglutaryl-CoA) reductase inhibitor, both intracellular FPP levels and PPARγ-target gene expressions were decreased. In contrast, the increase in intracellular FPP level after the addition of zaragozic acid, a squalene synthase inhibitor, induced PPARγ-target gene expression. The addition of FPP and zaragozic acid promotes lipid accumulation during adipocyte differentiation. These findings indicated that FPP might function as an endogenous PPARγ agonist and regulate gene expression in adipocytes.
© The Authors Journal compilation © 2011 Biochemical Society
Figures
Comment in
-
Isoprenoid is a perfect fit for fat factor.Biochem J. 2011 Aug 15;438(1):e1-3. doi: 10.1042/BJ20110996. Biochem J. 2011. PMID: 21793802
References
-
- Matsuzawa Y. The metabolic syndrome and adipocytokines. FEBS Lett. 2006;580:2917–2921. - PubMed
-
- Sakai T., Sakaue H., Nakamura T., Okada M., Matsuki Y., Watanabe E., Hiramatsu R., Nakayama K., Nakayama K. I., Kasuga M. Skp2 controls adipocyte proliferation during the development of obesity. J. Biol. Chem. 2007;282:2038–2046. - PubMed
-
- Spiegelman B. M., Flier J. S. Adipogenesis and obesity: rounding out the big picture. Cell. 1996;87:377–389. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous