

Review
doi: 10.1093/cvr/cvr138.
Epub 2011 May 23.
Epigenetic factors and cardiac development
Affiliations
- PMID: 21606181
- PMCID: PMC3125076
- DOI: 10.1093/cvr/cvr138
Item in Clipboard
Review
Epigenetic factors and cardiac development
Cardiovasc Res.
.
Abstract
Congenital heart malformations remain the leading cause of death related to birth defects. Recent advances in developmental and regenerative cardiology have shed light on a mechanistic understanding of heart development that is controlled by a transcriptional network of genetic and epigenetic factors. This article reviews the roles of chromatin remodelling factors important for cardiac development with the current knowledge of cardiac morphogenesis, regeneration, and direct cardiac differentiation. In the last 5 years, critical roles of epigenetic factors have been revealed in the cardiac research field.
Figures
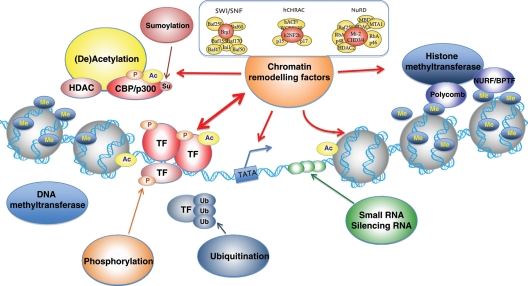
Chromatin structure and remodelling factors. DNA is organized in chromatin composed of condensed nucleosomes: units of 146 bp of DNA wrapped twice around an octamer of two copies of each histone protein H2A, H2B, H3, and H4. The flexible amino-terminal tails of the histones, protruding outward from the nucleosome, allow for post-translational modifications through (de)acethylation, phosporylation, ubiquitination, methylation, and sumoylation. Such covalent modifications alter DNA–histone interactions, affecting accessibility of transcription factors.
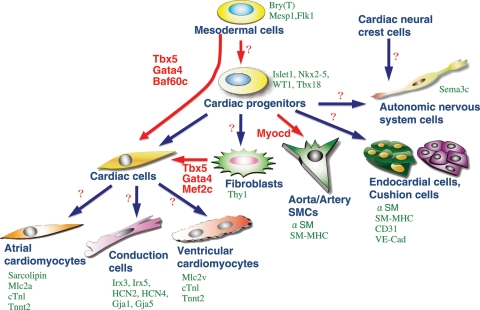
Cardiac cell types derived from multipotent progenitors. Differentiated cardiac cell types are marked by indicated genes (green). Recently, several factors have been defined as master regulators for cardiomyogenesis (red). The combination of Tbx5, Gata4, and Baf60c induces direct differentiation of mesodermal cells into ectopic beating myocytes, bypassing the cardiac progenitor state. Tbx5, Gata4, and Mef2c together can also induce cardiomyocytes from fibroblasts. Factors for direct induction of other cardiac cell types are currently unknown (question marks).
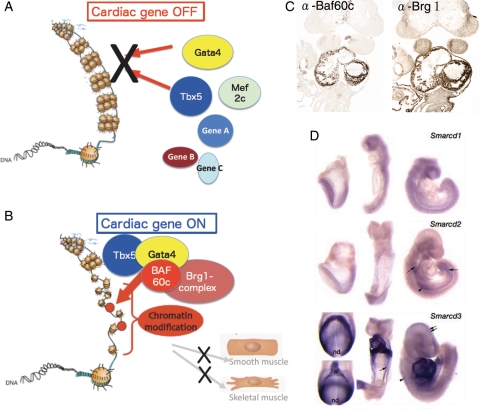
Chromatin remodelling factor-mediated regulation of cardiac transcription factors. (A) In the absence of Baf60c, the cardiac transcription factors Gata4, Mef2c, and Tbx5 may not have access to their target genes. The highly condensed euchromatin in which the DNA is tightly wrapped around histones makes transcription factors inaccessible to regulatory DNA, thereby represses transcription. (B) Chromatin remodelling factors modify chromatin organization by unwinding DNA from the histones, making target sequences accessible for transcription factors. Baf60c acts as a bridge to bring the Brg1 complex together with the transcription factors Tbx5 and Gata4 in a tissue-specific manner. (C) Heart-restricted expression of Baf60c and Brg1 shown by immunostaining. (D) Expression of Smarcd3 (encoding Baf60c) restricted to cardiogenic mesoderm and the developing heart shown by in situ hybridization (adapted from Lickert et al.).
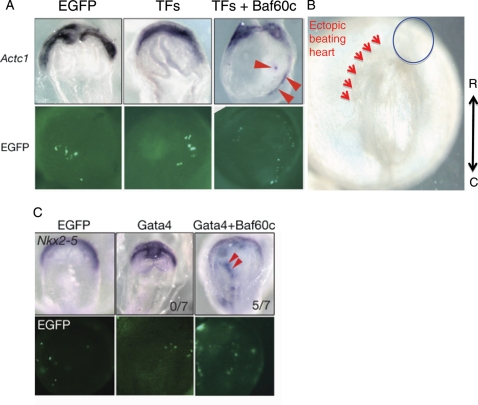
Ectopic induction of cardiomyogenesis by defined factors (Tbx5, Gata4, and Baf60c). (A) Ectopic induction of cardiac tissues by co-overexpression of TFs (Tbx5, Nkx2–5, and Gata4) and Baf60c. The early cardiomyocyte marker Actc1 was used to monitor the induction of cardiomyocytes. The chromatin remodelling component Baf60c is required for the induction. (B) Beating heart tissues (arrowheads) are observed in non-cardiogenic mesoderm upon overexpression of Tbx5, Gata4, and Baf60c. At this stage, the endogenous heart cells do not beat, indicating accelerated cardiac differentiation by the defined factors. (C) Whole-mount in situ hybridization showing that Gata4 requires Baf60c to induce ectopic expression of Nkx2–5. EGFP expression indicates transfected cells (adapted from Takeuchi and Bruneau).
References
-
- Abu-Issa R, Kirby ML. Heart field: from mesoderm to heart tube. Annu Rev Cell Dev Biol. 2007;23:45–68. doi:10.1146/annurev.cellbio.23.090506.123331. - DOI - PubMed
-
- Buckingham M, Meilhac S, Zaffran S. Building the mammalian heart from two sources of myocardial cells. Nat Rev Genet. 2005;6:826–835. doi:10.1038/nrg1710. - DOI - PubMed
-
- Garry DJ, Olson EN. A common progenitor at the heart of development. Cell. 2006;127:1101–1104. doi:10.1016/j.cell.2006.11.031. - DOI - PubMed
-
- Moorman AFM, Webb S, Brown NA, Lamers W, Anderson RH. Development of the heart: (1) formation of the cardiac chambers and arterial trunks. Heart. 2003;89:806–814. doi:10.1136/heart.89.7.806. - DOI - PMC - PubMed
-
- Christoffels VM, Habets PE, Franco D, Campione M, de Jong F, Lamers WH, et al. Chamber formation and morphogenesis in the developing mammalian heart. Dev Biol. 2000;223:266–278. doi:10.1006/dbio.2000.9753. - DOI - PubMed
