

IL-6 mediates the susceptibility of glycoprotein 130 hypermorphs to Toxoplasma gondii
- PMID: 21606248
- PMCID: PMC3119722
- DOI: 10.4049/jimmunol.1004144
IL-6 mediates the susceptibility of glycoprotein 130 hypermorphs to Toxoplasma gondii
Abstract
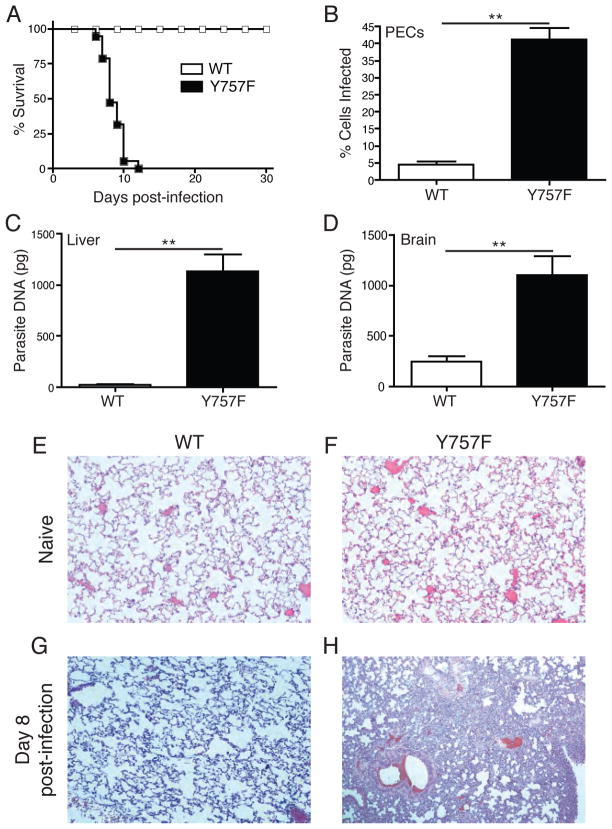
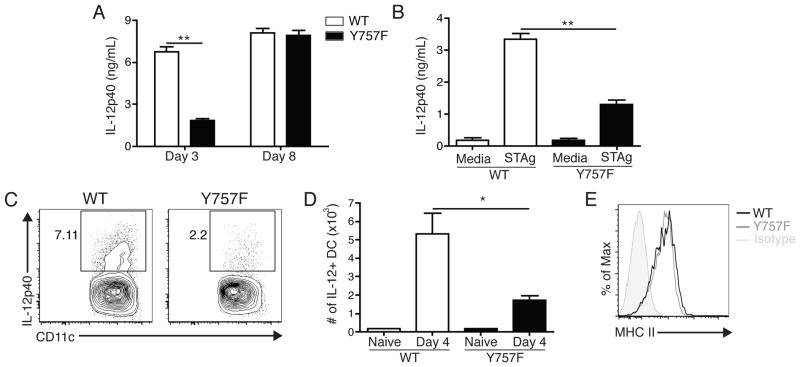
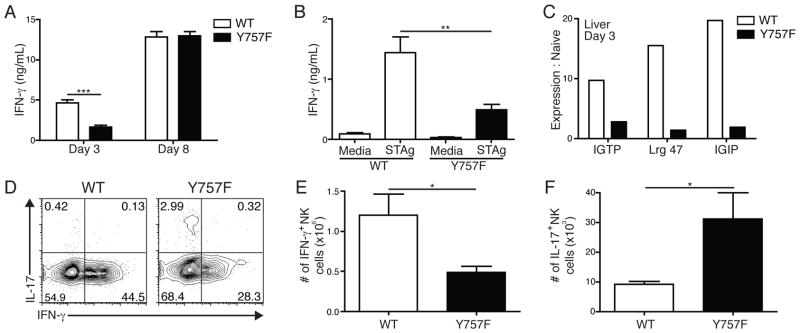
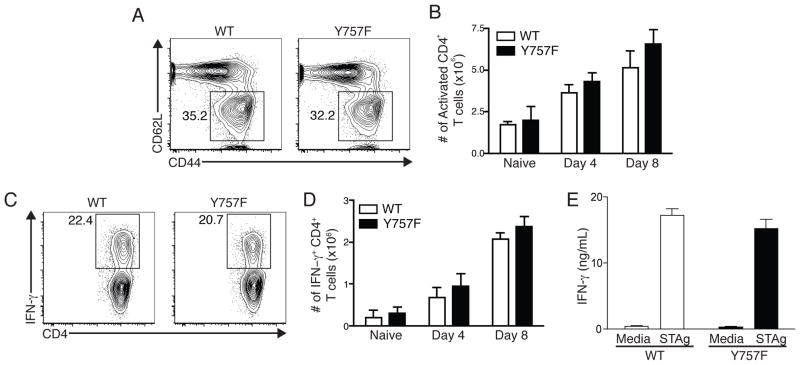
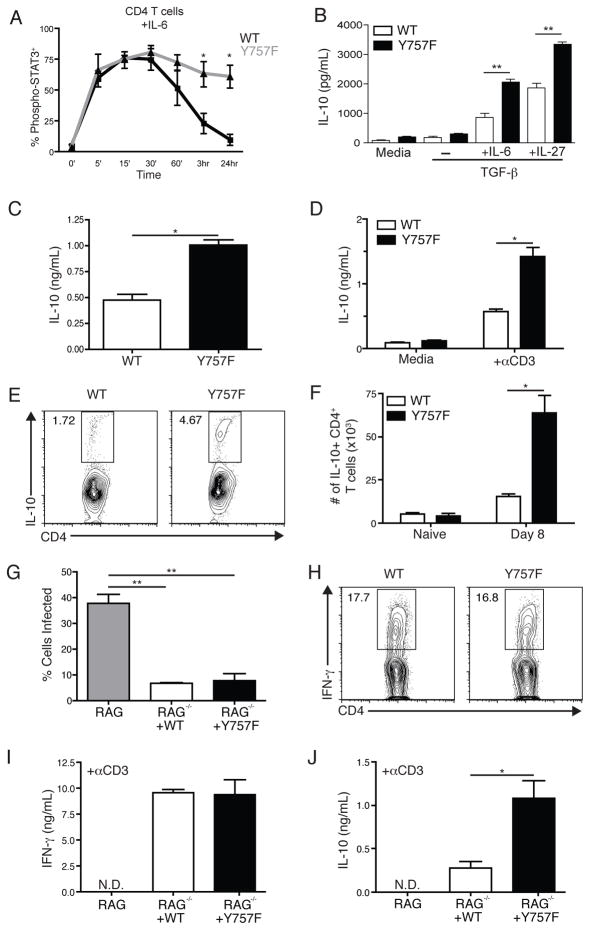
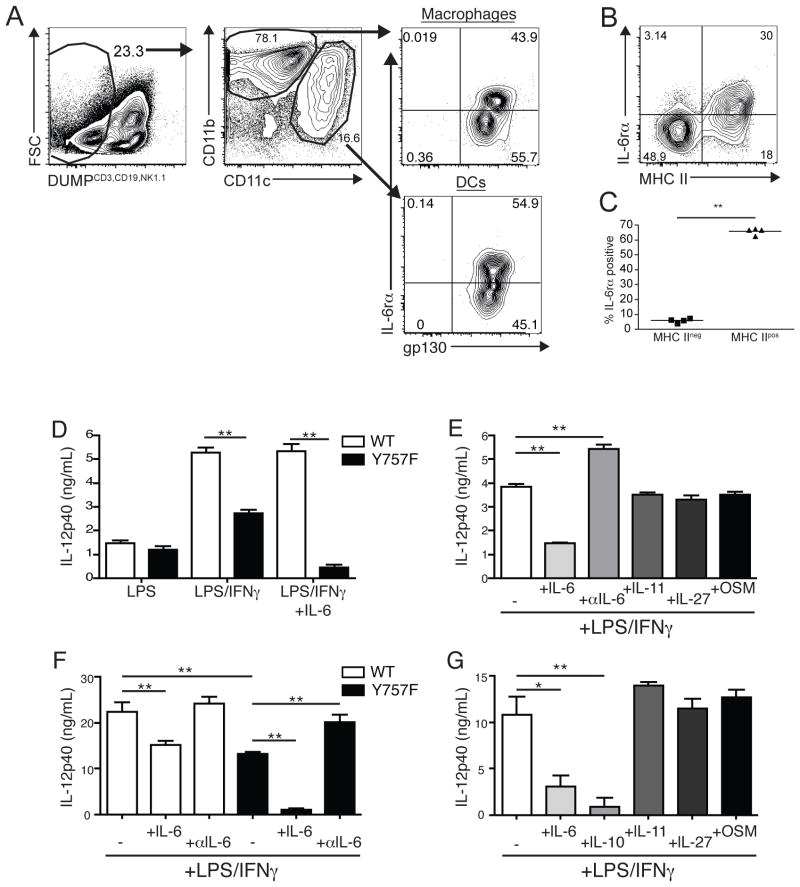
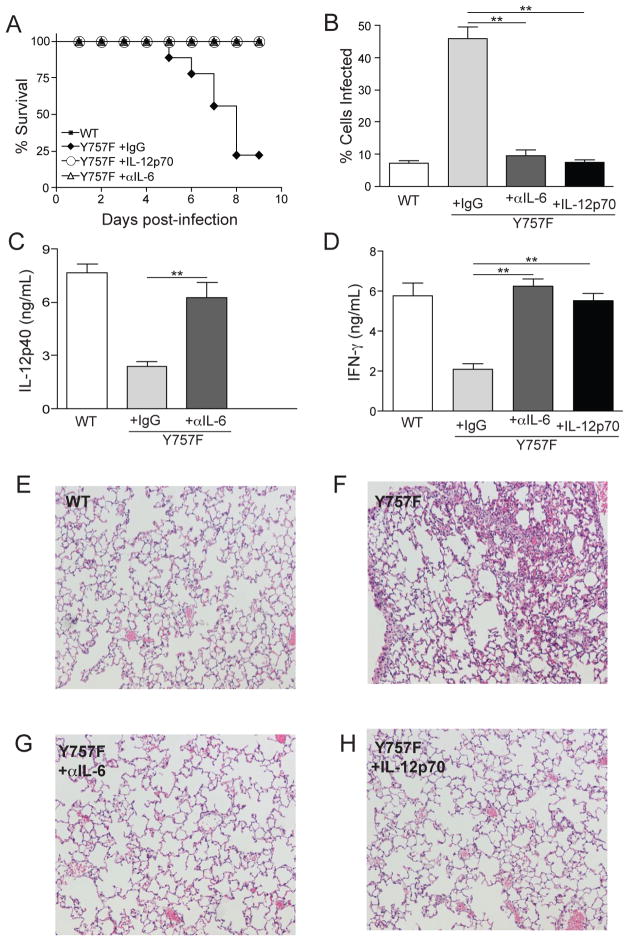
IL-6 and IL-27 are closely related cytokines that play critical but distinct roles during infection with Toxoplasma gondii. Thus, IL-6 is required for the development of protective immunity to this pathogen, whereas IL-27 is required to limit infection-induced pathology. Paradoxically, these factors both signal through gp130, but little is known about how the signals downstream of gp130 are integrated to coordinate the immune response to infection. To better understand these events, gp130 Y757F mice that have a mutation in gp130 at the binding site for suppressor of cytokine signaling 3, a critical negative regulator of gp130 signaling, were infected with T. gondii. These mutant mice were acutely susceptible to this challenge, characterized by an early defect in the production of IL-12 and IFN-γ and increased parasite burdens. Consistent with the reduced IL-12 levels, IL-6, but not other gp130 cytokines, was a potent antagonist of IL-12 production by gp130 Y757F macrophages and dendritic cells in vitro. Moreover, in gp130 Y757F mice, blocking IL-6 in vivo, or administration of rIL-12, during infection restored IFN-γ production and protective immunity. Collectively, these studies highlight that a failure to abbreviate IL-6-mediated gp130 signaling results in a profound anti-inflammatory signal that blocks the generation of protective immunity to T. gondii.
Figures
References
-
- Taga T, Kishimoto T. Gp130 and the interleukin-6 family of cytokines. Annu Rev Immunol. 1997;15:797–819. - PubMed
-
- O’Shea JJ, Gadina M, Schreiber RD. Cytokine signaling in 2002: new surprises in the Jak/Stat pathway. Cell. 2002;109(Suppl):S121–31. - PubMed
-
- De Souza D, Fabri LJ, Nash A, Hilton DJ, Nicola NA, Baca M. SH2 domains from suppressor of cytokine signaling-3 and protein tyrosine phosphatase SHP-2 have similar binding specificities. Biochemistry. 2002;41:9229–9236. - PubMed
-
- Nicholson SE, De Souza D, Fabri LJ, Corbin J, Willson TA, Zhang JG, Silva A, Asimakis M, Farley A, Nash AD, Metcalf D, Hilton DJ, Nicola NA, Baca M. Suppressor of cytokine signaling-3 preferentially binds to the SHP-2-binding site on the shared cytokine receptor subunit gp130. Proc Natl Acad Sci U S A. 2000;97:6493–6498. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
