

Recognition of the centromere-specific histone Cse4 by the chaperone Scm3
- PMID: 21606327
- PMCID: PMC3111289
- DOI: 10.1073/pnas.1106389108
Recognition of the centromere-specific histone Cse4 by the chaperone Scm3
Abstract
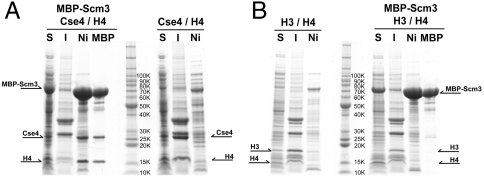
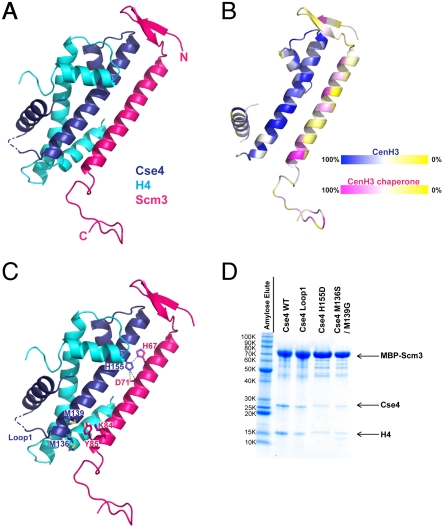
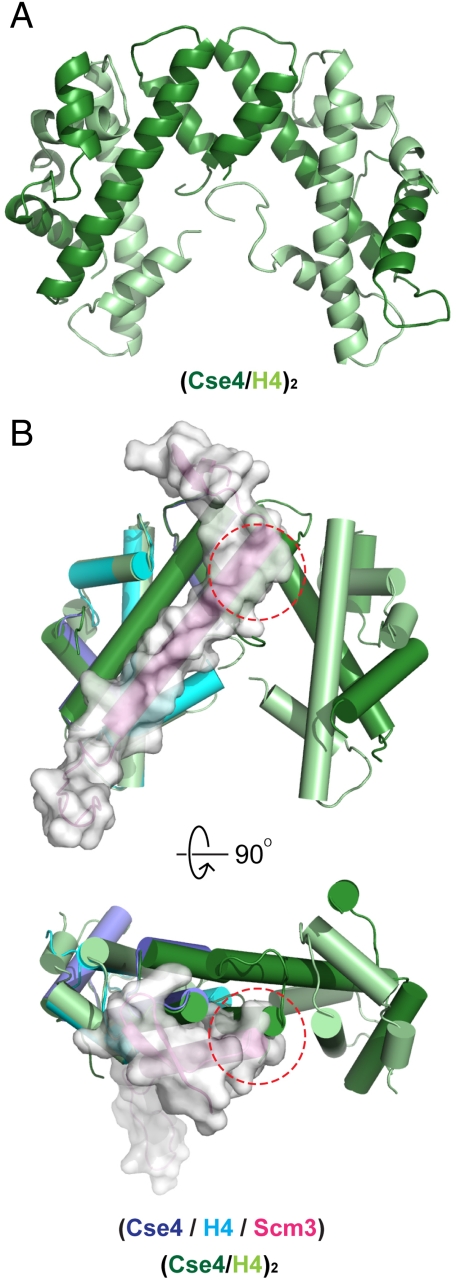
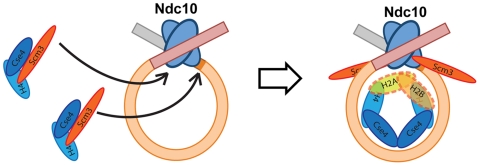
A specialized nucleosome is a component of all eukaryotic kinetochores. The core of this nucleosome contains a centromere-specific histone, CENP-A (the Cse4 gene product in budding yeast), instead of the usual H3. Assembly of a centromeric nucleosome depends on a specific chaperone, called Scm3 in yeast and HJURP in higher eukaryotes. We describe here the structure of a complex formed by an N-terminal fragment of Scm3 with the histone-fold domains of Cse4, and H4, all prepared as recombinant proteins derived from the budding yeast Kluyveromyces lactis. The contacts of Scm3 with Cse4 explain its selectivity for the centromere-specific histone; key residues at the interface are conserved in HJURP, indicating a common mechanism for centromeric-histone deposition. We also report the structure of a (Cse4 : H4)(2) heterotetramer; comparison with the structure of the Scm3:Cse4:H4 complex shows that tetramer formation and DNA-binding require displacement of Scm3 from the nucleosome core. The two structures together suggest that specific contacts between the chaperone and Cse4, rather than an altered overall structure of the nucleosome core, determine the selective presence of Cse4 at centromeres.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Structure of the budding yeast Saccharomyces cerevisiae centromeric histones Cse4-H4 complexed with the chaperone Scm3.Proc Natl Acad Sci U S A. 2011 Aug 30;108(35):E596; author reply E597. doi: 10.1073/pnas.1109548108. Epub 2011 Aug 15. Proc Natl Acad Sci U S A. 2011. PMID: 21844330 Free PMC article. No abstract available.
References
-
- McAinsh AD, Tytell JD, Sorger PK. Structure, function, and regulation of budding yeast kinetochores. Annu Rev Cell Dev Biol. 2003;19:519–539. - PubMed
-
- Earnshaw WC, Rothfield N. Identification of a family of human centromere proteins using autoimmune sera from patients with scleroderma. Chromosoma. 1985;91:313–321. - PubMed
-
- Meluh PB, Yang P, Glowczewski L, Koshland D, Smith MM. Cse4p is a component of the core centromere of Saccharomyces cerevisiae. Cell. 1998;94:607–613. - PubMed
-
- Stoler S, Keith KC, Curnick KE, Fitzgerald-Hayes M. A mutation in CSE4, an essential gene encoding a novel chromatin-associated protein in yeast, causes chromosome nondisjunction and cell cycle arrest at mitosis. Genes Dev. 1995;9:573–586. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
