

Systematic variation in the temperature dependence of physiological and ecological traits
- PMID: 21606358
- PMCID: PMC3127911
- DOI: 10.1073/pnas.1015178108
Systematic variation in the temperature dependence of physiological and ecological traits
Abstract
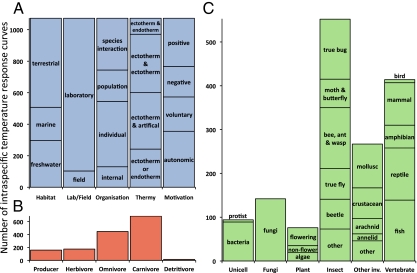
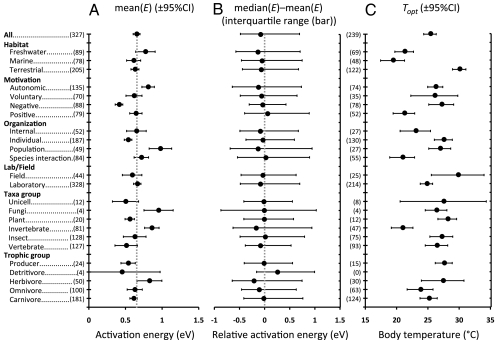
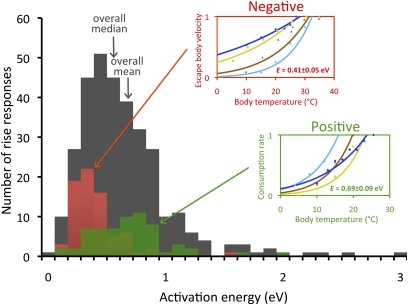
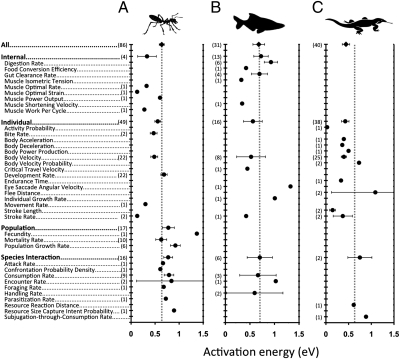
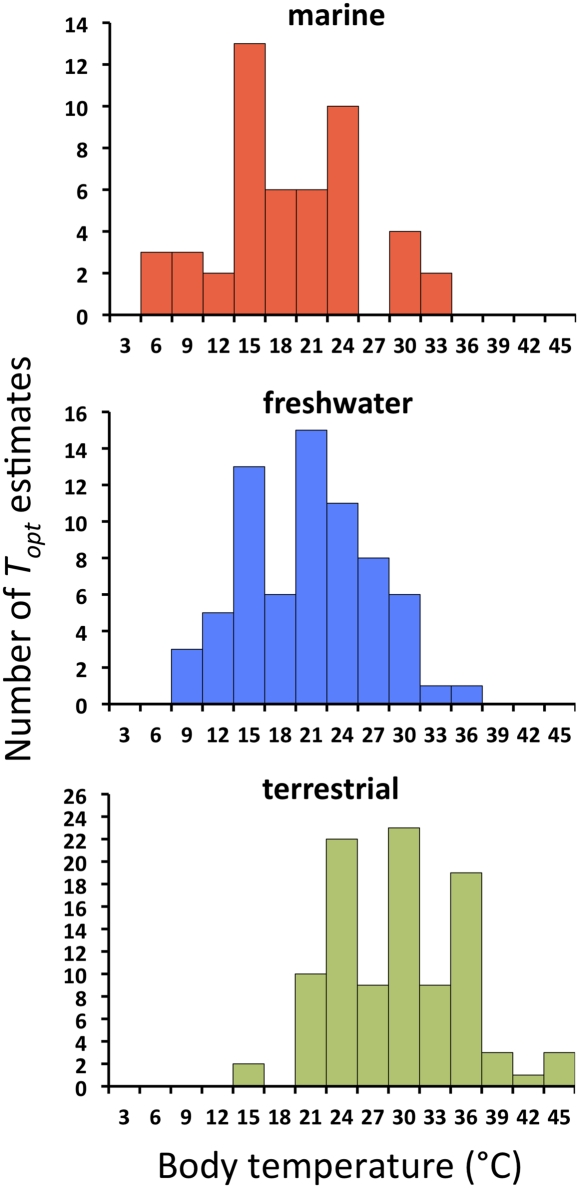
To understand the effects of temperature on biological systems, we compile, organize, and analyze a database of 1,072 thermal responses for microbes, plants, and animals. The unprecedented diversity of traits (n = 112), species (n = 309), body sizes (15 orders of magnitude), and habitats (all major biomes) in our database allows us to quantify novel features of the temperature response of biological traits. In particular, analysis of the rising component of within-species (intraspecific) responses reveals that 87% are fit well by the Boltzmann-Arrhenius model. The mean activation energy for these rises is 0.66 ± 0.05 eV, similar to the reported across-species (interspecific) value of 0.65 eV. However, systematic variation in the distribution of rise activation energies is evident, including previously unrecognized right skewness around a median of 0.55 eV. This skewness exists across levels of organization, taxa, trophic groups, and habitats, and it is partially explained by prey having increased trait performance at lower temperatures relative to predators, suggesting a thermal version of the life-dinner principle-stronger selection on running for your life than running for your dinner. For unimodal responses, habitat (marine, freshwater, and terrestrial) largely explains the mean temperature at which trait values are optimal but not variation around the mean. The distribution of activation energies for trait falls has a mean of 1.15 ± 0.39 eV (significantly higher than rises) and is also right-skewed. Our results highlight generalities and deviations in the thermal response of biological traits and help to provide a basis to predict better how biological systems, from cells to communities, respond to temperature change.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Variation in universal temperature dependence of biological rates.Proc Natl Acad Sci U S A. 2011 Jun 28;108(26):10377-8. doi: 10.1073/pnas.1107430108. Epub 2011 Jun 16. Proc Natl Acad Sci U S A. 2011. PMID: 21680885 Free PMC article. No abstract available.
References
-
- IPCC. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland: IPPC; 2007.
-
- Brown JH, Gillooly JF, Allen AP, Savage VM, West GB. Toward a metabolic theory of ecology. Ecology. 2004;85:1771–1789.
-
- Angilletta MJ. Thermal adaptation: A Theoretical and Empirical Synthesis. Oxford: Oxford Univ Press; 2009.
-
- Gillooly JF, Brown JH, West GB, Savage VM, Charnov EL. Effects of size and temperature on metabolic rate. Science. 2001;293:2248–2251. - PubMed
-
- Cossins AR, Bowler K. Temperature Biology of Animals. New York: Chapman & Hall; 1987.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
