

Retinal determination genes function along with cell-cell signals to regulate Drosophila eye development: examples of multi-layered regulation by master regulators
- PMID: 21607995
- PMCID: PMC4300108
- DOI: 10.1002/bies.201000131
Retinal determination genes function along with cell-cell signals to regulate Drosophila eye development: examples of multi-layered regulation by master regulators
Abstract
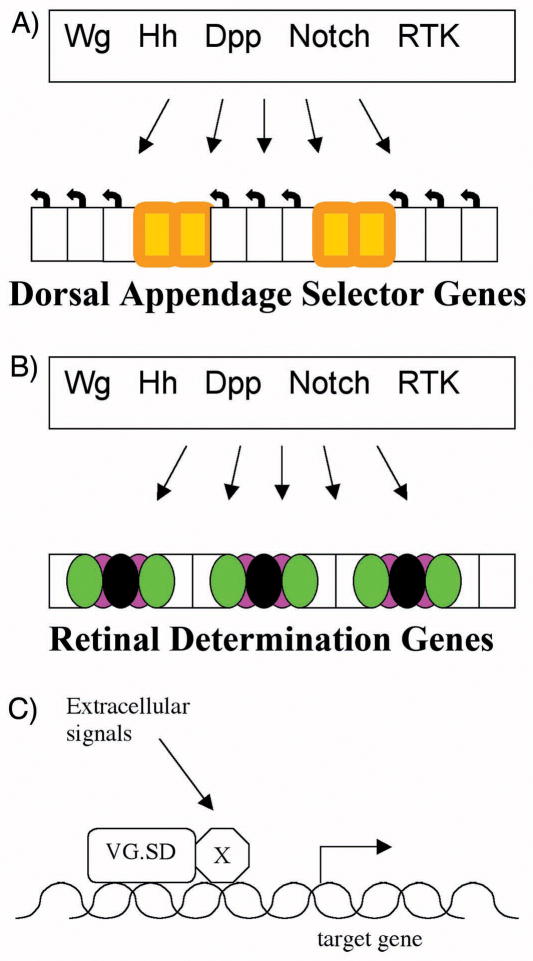
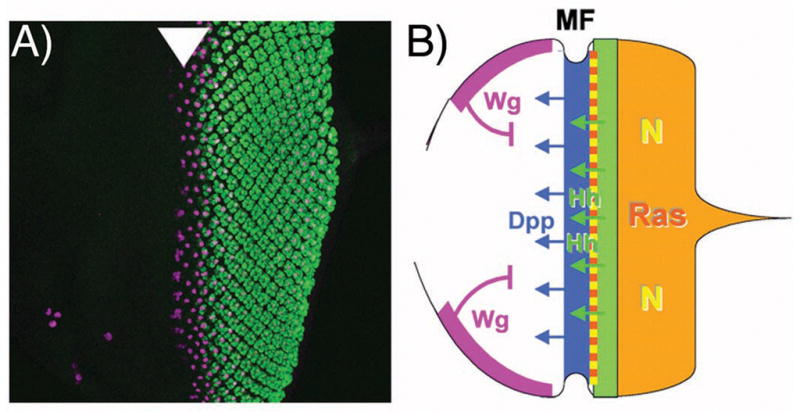
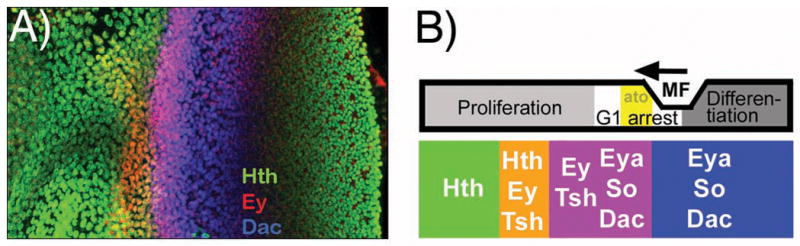
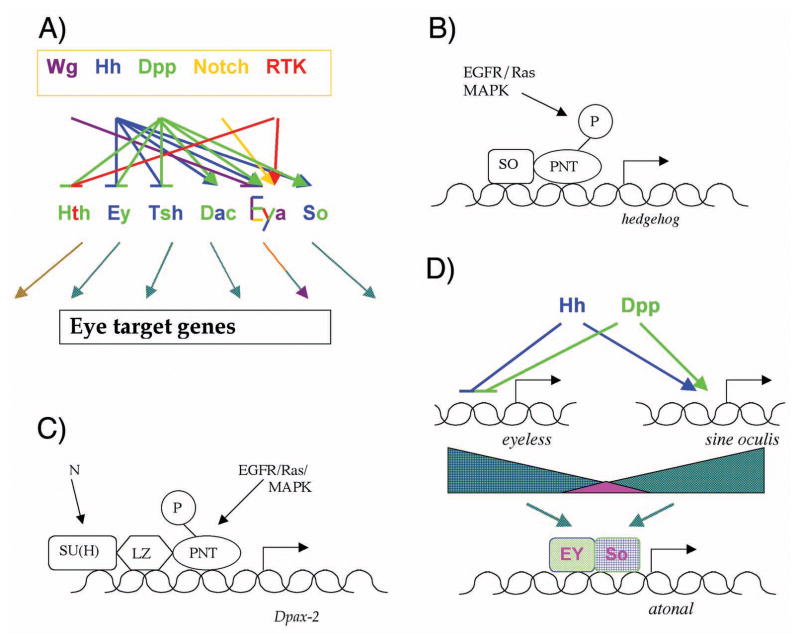
It is thought that retinal determination (RD) gene products define the response made to cell-cell signals in the field of eye development by binding to enhancers of genes that are also regulated by cell-cell signaling pathways. In Drosophila, RD genes, including eyeless, teashirt, eyes absent, dachsous, and sine oculis, are required for normal eye development and can induce ectopic eyes when mis-expressed. Characterization of the enhancers responsible for eye expression of the hedgehog, shaven, and atonal genes, as well as the dynamics of RD gene expression themselves, now suggest a multilayered network whereby transcriptional regulation by either RD genes or cell-cell signaling pathways can sometimes be indirect and mediated by other transcription factor intermediates. In this updated view of the interaction between extracellular information and cell intrinsic programs during development, regulation of individual genes might sometimes be several steps removed from either the RD genes or the cell-cell signaling pathways that nevertheless govern their expression.
Copyright © 2011 WILEY Periodicals, Inc.
Figures
References
-
- Halder G, Callaerts P, Gehring WJ. Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science. 1995;267:1788–92. - PubMed
-
- Curtiss J, Halder G, Mlodzik M. Selector and signalling molecules cooperate in organ patterning. Nat Cell Biol. 2002;4:E48–51. - PubMed
-
- Mann RS, Carroll SB. Molecular mechanisms of selector gene function and evolution. Curr Opin Genet Dev. 2002;12:592–600. - PubMed
-
- Gerhart J. Warkany lecture: signaling pathways in development. Teratology. 1999;60:226–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
