

A photoreactive small-molecule probe for 2-oxoglutarate oxygenases
- PMID: 21609845
- PMCID: PMC4681097
- DOI: 10.1016/j.chembiol.2011.03.007
A photoreactive small-molecule probe for 2-oxoglutarate oxygenases
Abstract
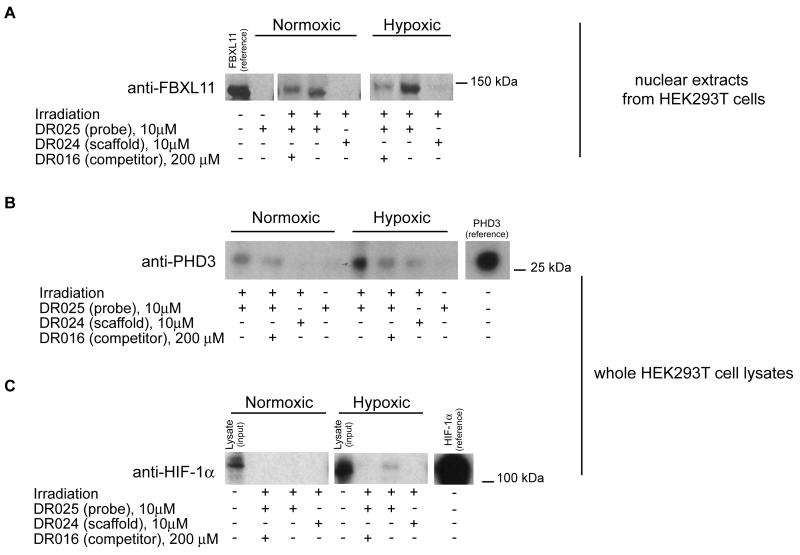
2-oxoglutarate (2-OG)-dependent oxygenases have diverse roles in human biology. The inhibition of several 2-OG oxygenases is being targeted for therapeutic intervention, including for cancer, anemia, and ischemic diseases. We report a small-molecule probe for 2-OG oxygenases that employs a hydroxyquinoline template coupled to a photoactivable crosslinking group and an affinity-purification tag. Following studies with recombinant proteins, the probe was shown to crosslink to 2-OG oxygenases in human crude cell extracts, including to proteins at endogenous levels. This approach is useful for inhibitor profiling, as demonstrated by crosslinking to the histone demethylase FBXL11 (KDM2A) in HEK293T nuclear extracts. The results also suggest that small-molecule probes may be suitable for substrate identification studies.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
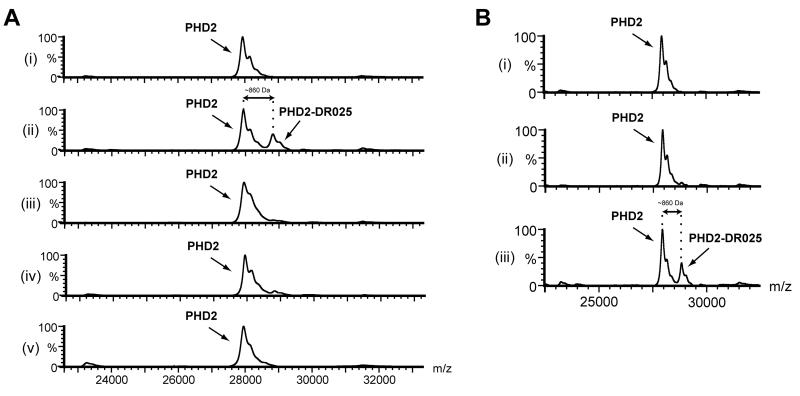
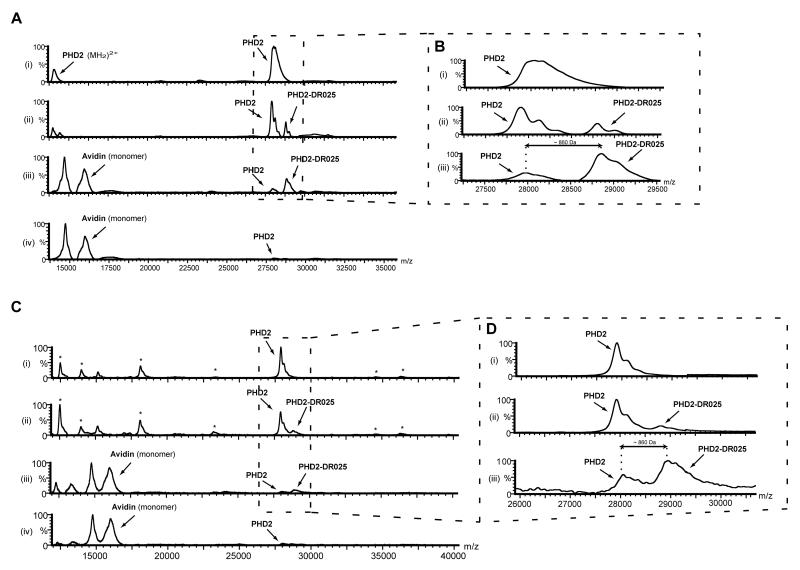
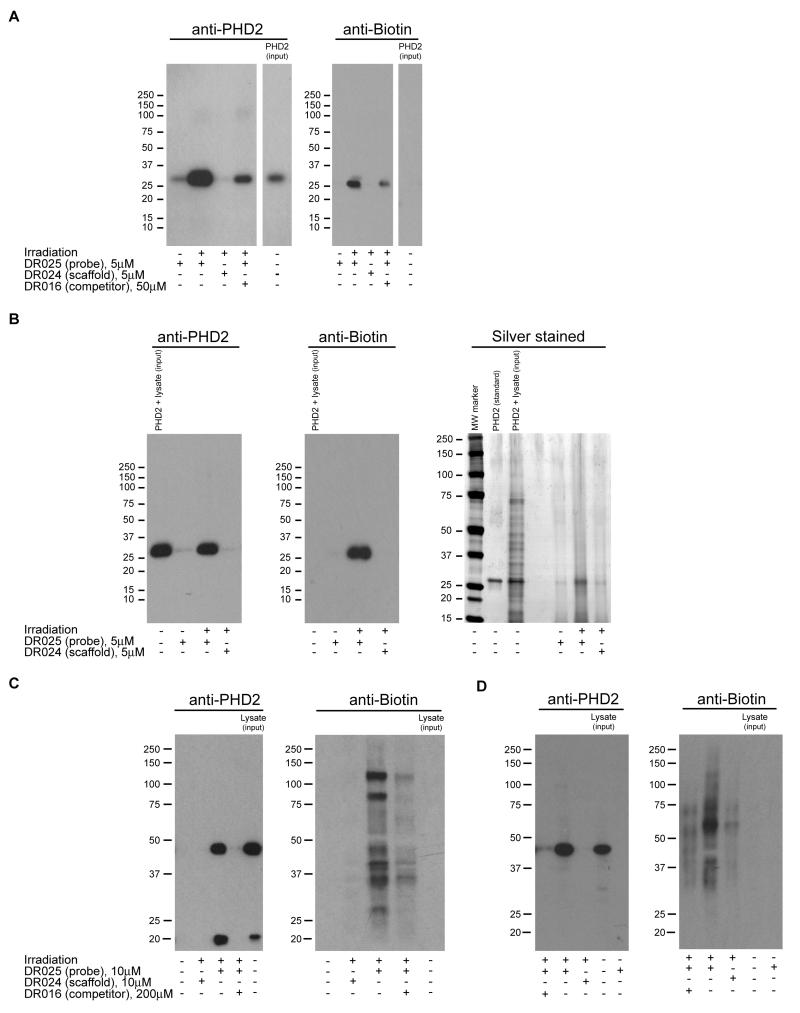
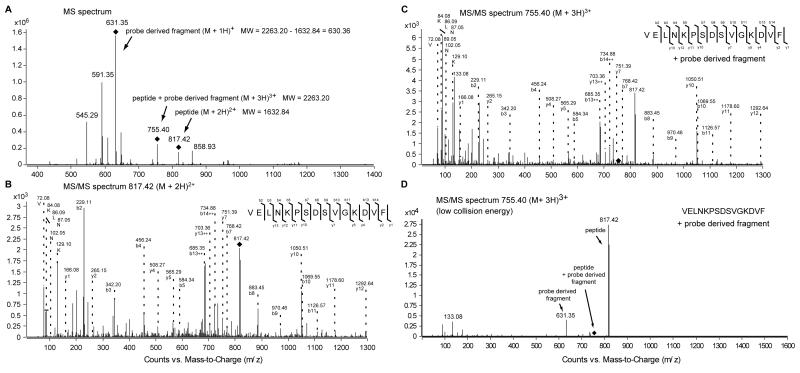
Comment in
-
Activity-based profiling of 2-oxoglutarate oxygenases.Chem Biol. 2011 May 27;18(5):557-9. doi: 10.1016/j.chembiol.2011.05.002. Chem Biol. 2011. PMID: 21609835
References
-
- Appelhoff RJ, Tian YM, Raval RR, Turley H, Harris AL, Pugh CW, Ratcliffe PJ, Gleadle JM. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J. Biol. Chem. 2004;279:38458–38465. - PubMed
-
- Chowdhury R, McDonough MA, Mecinovic J, Loenarz C, Flashman E, Hewitson KS, Domene C, Schofield CJ. Structural basis for binding of hypoxia-inducible factor to the oxygen-sensing prolyl hydroxylases. Structure. 2009;17:981–989. - PubMed
-
- Epstein ACR, Gleadle JM, McNeill LA, Hewitson KS, O’Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, et al. C. elegans EGL-9 and Mammalian Homologs Define a Family of Dioxygenases that Regulate HIF by Prolyl Hydroxylation. Cell. 2001;107:43–54. - PubMed
-
- Evans MJ, Cravatt BF. Mechanism-based profiling of enzyme families. Chem. Rev. 2006;106:3279–3301. - PubMed
-
- Fisher JJ, Graebner(nee’ Baessler) OY, Dalhoff C, Michaelis S, Schrey AK, Ungewiss J, Andrich K, Jeske D, Kroll F, Glinski M, et al. Comprehensive Identification of Staurosporine-Binding Kinases in the Hepatocyte Cell Line HepG2 using Capture Compound Mass Spectrometry (CCMS) J. Proteome Res. 2010;9:806–817. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
