

An interaction between the Walker A and D-loop motifs is critical to ATP hydrolysis and cooperativity in bacteriophage T4 Rad50
- PMID: 21610075
- PMCID: PMC3138309
- DOI: 10.1074/jbc.M111.256305
An interaction between the Walker A and D-loop motifs is critical to ATP hydrolysis and cooperativity in bacteriophage T4 Rad50
Abstract
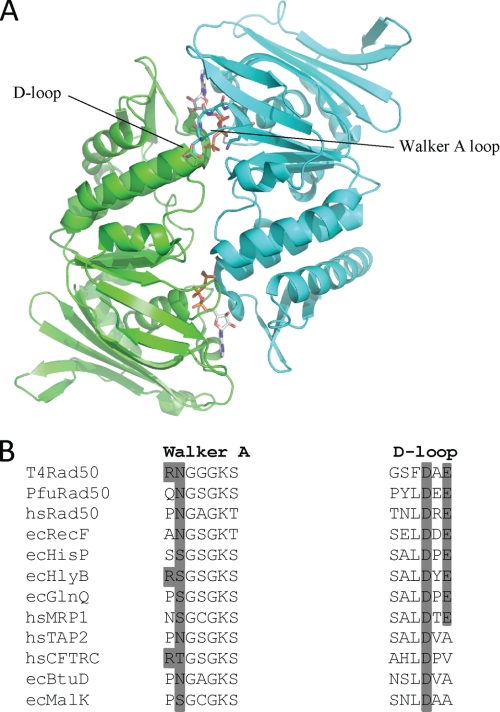
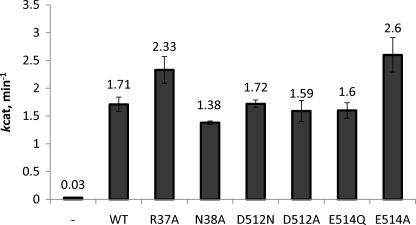
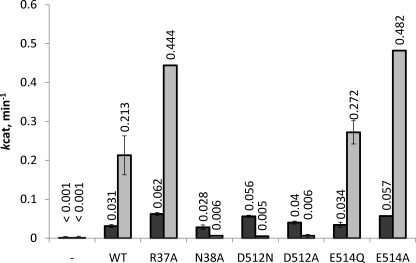
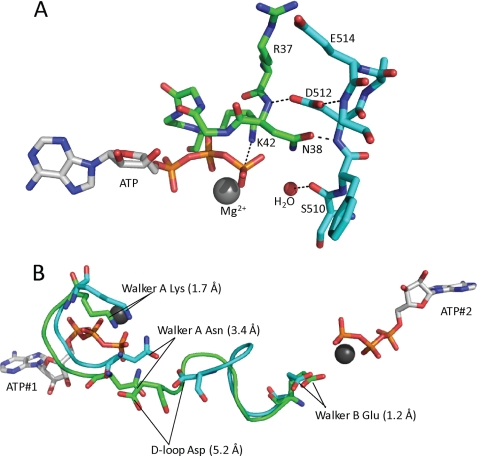
The ATP binding cassette (ABC) proteins make up a large superfamily with members coming from all kingdoms. The functional form of the ABC protein nucleotide binding domain (NBD) is dimeric with ATP binding sites shared between subunits. The NBD is defined by six motifs: the Walker A, Q-loop, Signature, Walker-B, D-loop, and H-loop. The D-loop contains a conserved aspartate whose function is not clear but has been proposed to be involved in cross-talk between ATP binding sites. Structures of various ABC proteins suggest an interaction between the D-loop aspartate and an asparagine residue located in Walker A loop of the opposing subunit. Here, we evaluate the functional role of the D-loop using a bacteriophage T4 ABC protein, Rad50 (gp46). Mutation of either the D-loop aspartate or the Walker A asparagine results in dramatic reductions in ATP affinity, hydrolysis rate, and cooperativity. The mutant proteins bind Mre11 (gp47) and DNA normally, but no longer support the ATP-dependent nuclease activities of Mre11. We propose that the D-loop aspartate functions to stabilize the Walker A asparagine in a position favorable for catalysis. We find that the asparagine is crucially important to the mechanism of ATP hydrolysis by increasing the affinity for ATP and positioning the γ-phosphate of ATP for catalysis. Additionally, we propose that the asparagine acts as a γ-phosphate sensor and, through its interaction with the conserved D-loop aspartate, transmits conformational changes across the dimer interface to the second ATP binding site.
Figures
References
-
- Connelly J. C., Leach D. R. (2002) Trends Biochem. Sci. 27, 410–418 - PubMed
-
- Aguilera A., Gómez-González B. (2008) Nat. Rev. Genet. 9, 204–217 - PubMed
-
- Hoeijmakers J. H. J. (2007) Mech. Ageing Dev. 128, 460–462 - PubMed
-
- Borde V. (2007) Chromosome Res. 15, 551–563 - PubMed
-
- Soulas-Sprauel P., Rivera-Munoz P., Malivert L., Le Guyader G., Abramowski V., Revy P., de Villartay J. (2007) Oncogene 26, 7780–7791 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
