

HOPS drives vacuole fusion by binding the vacuolar SNARE complex and the Vam7 PX domain via two distinct sites
- PMID: 21613544
- PMCID: PMC3135484
- DOI: 10.1091/mbc.E11-02-0104
HOPS drives vacuole fusion by binding the vacuolar SNARE complex and the Vam7 PX domain via two distinct sites
Abstract
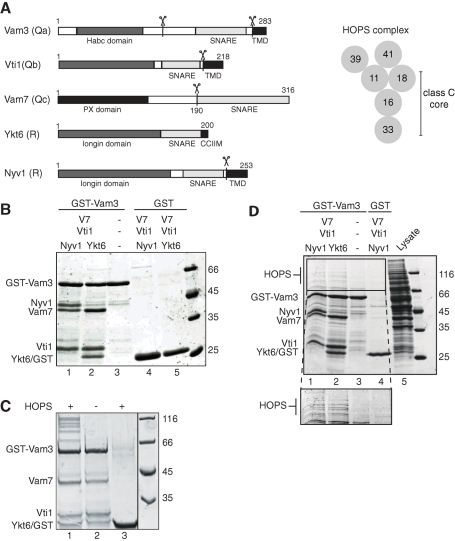
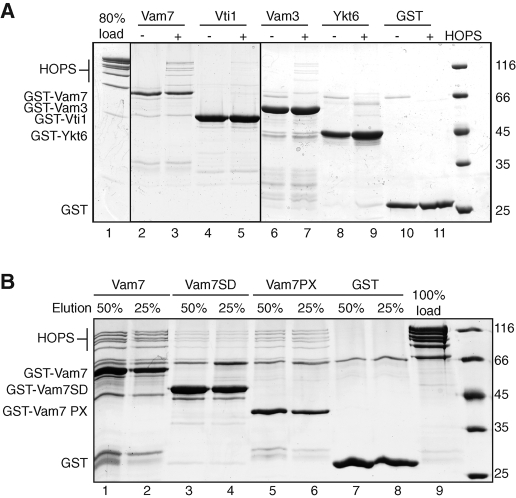
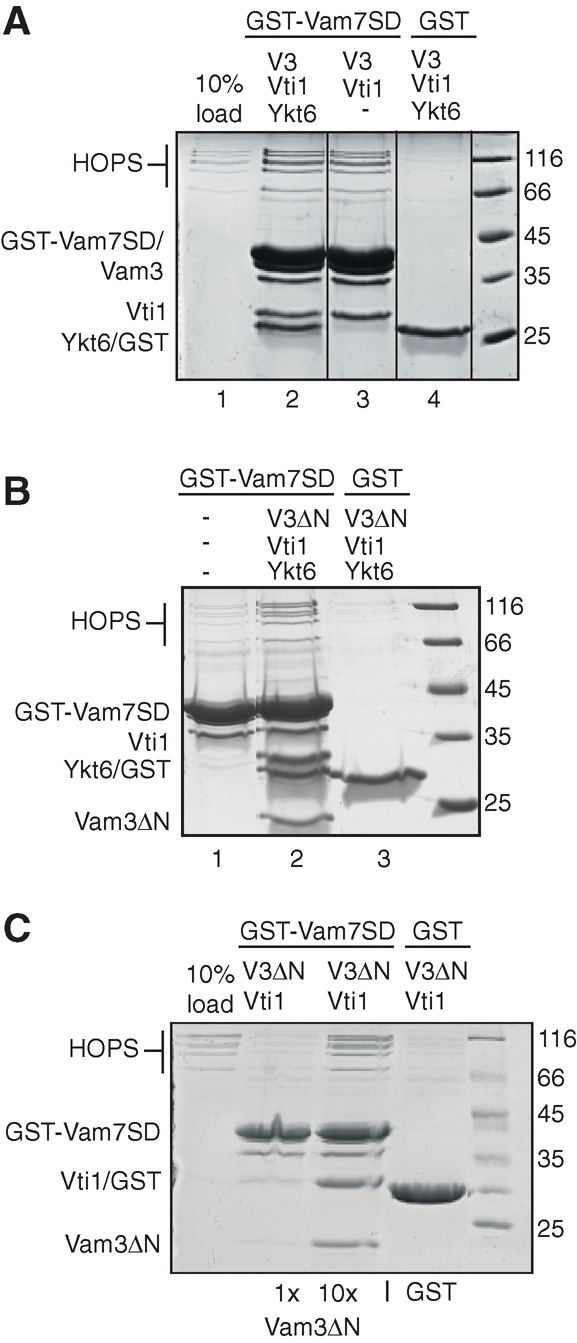
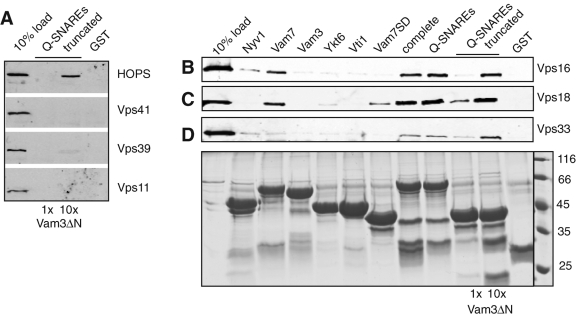
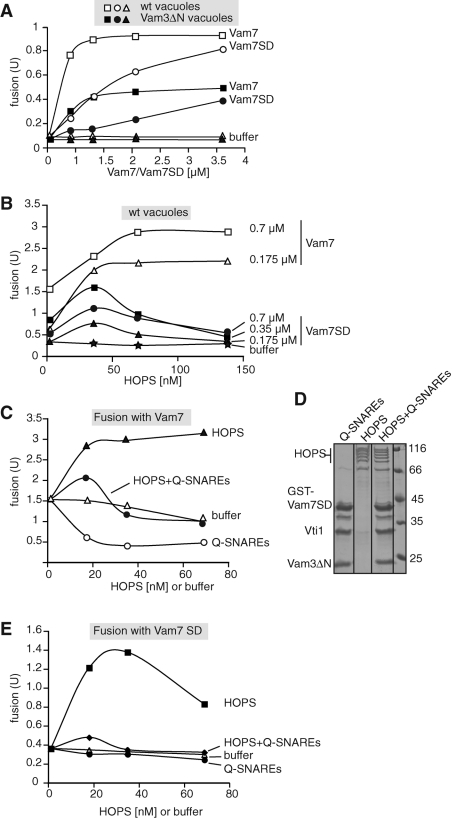
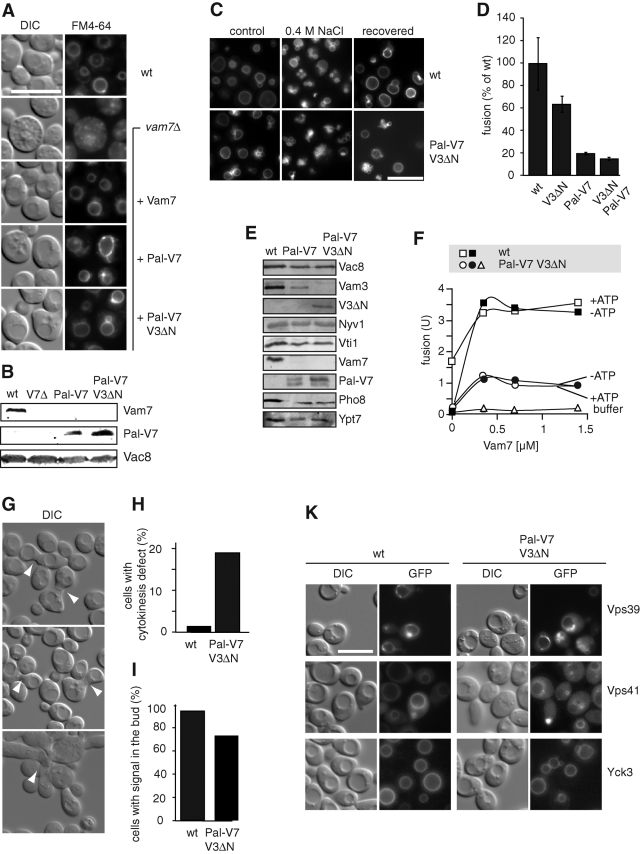
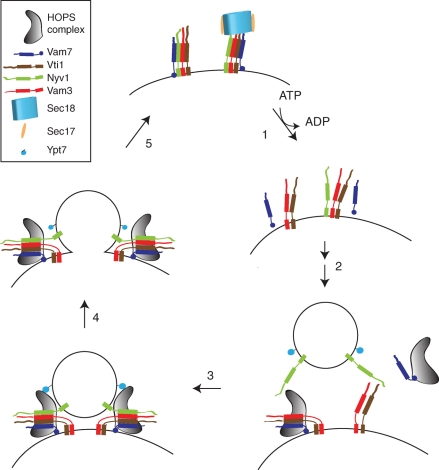
Membrane fusion within the endomembrane system follows a defined order of events: membrane tethering, mediated by Rabs and tethers, assembly of soluble N-ethylmaleimide-sensitive factor (NSF) attachment protein receptor (SNARE) complexes, and lipid bilayer mixing. Here we present evidence that the vacuolar HOPS tethering complex controls fusion through specific interactions with the vacuolar SNARE complex (consisting of Vam3, Vam7, Vti1, and Nyv1) and the N-terminal domains of Vam7 and Vam3. We show that homotypic fusion and protein sorting (HOPS) binds Vam7 via its subunits Vps16 and Vps18. In addition, we observed that Vps16, Vps18, and the Sec1/Munc18 protein Vps33, which is also part of the HOPS complex, bind to the Q-SNARE complex. In agreement with this observation, HOPS-stimulated fusion was inhibited if HOPS was preincubated with the minimal Q-SNARE complex. Importantly, artificial targeting of Vam7 without its PX domain to membranes rescued vacuole morphology in vivo, but resulted in a cytokinesis defect if the N-terminal domain of Vam3 was also removed. Our data thus support a model of HOPS-controlled membrane fusion by recognizing different elements of the SNARE complex.
Figures
References
-
- Bowers K, Stevens TH. Protein transport from the late Golgi to the vacuole in the yeast Saccharomyces cerevisiae. Biochim Biophys Acta. 2005;1744:438–454. - PubMed
-
- Brachmann CB, Davies A, Cost GJ, Caputo E, Li J, Hieter P, Boeke JD. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast. 1998;14:115–132. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
