

Behavioral epigenetics
- PMID: 21615751
- PMCID: PMC3783959
- DOI: 10.1111/j.1749-6632.2011.06037.x
Behavioral epigenetics
Abstract
Sponsored by the New York Academy of Sciences, the Warren Alpert Medical School of Brown University and the University of Massachusetts Boston, "Behavioral Epigenetics" was held on October 29-30, 2010 at the University of Massachusetts Boston Campus Center, Boston, Massachusetts. This meeting featured speakers and panel discussions exploring the emerging field of behavioral epigenetics, from basic biochemical and cellular mechanisms to the epigenetic modulation of normative development, developmental disorders, and psychopathology. This report provides an overview of the research presented by leading scientists and lively discussion about the future of investigation at the behavioral epigenetic level.
© 2011 New York Academy of Sciences.
Figures
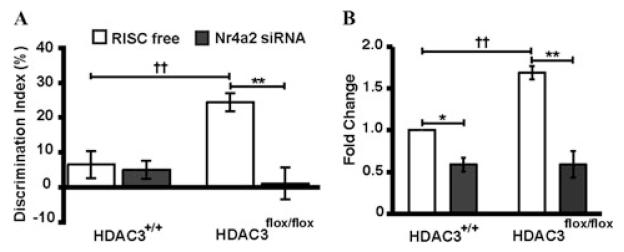
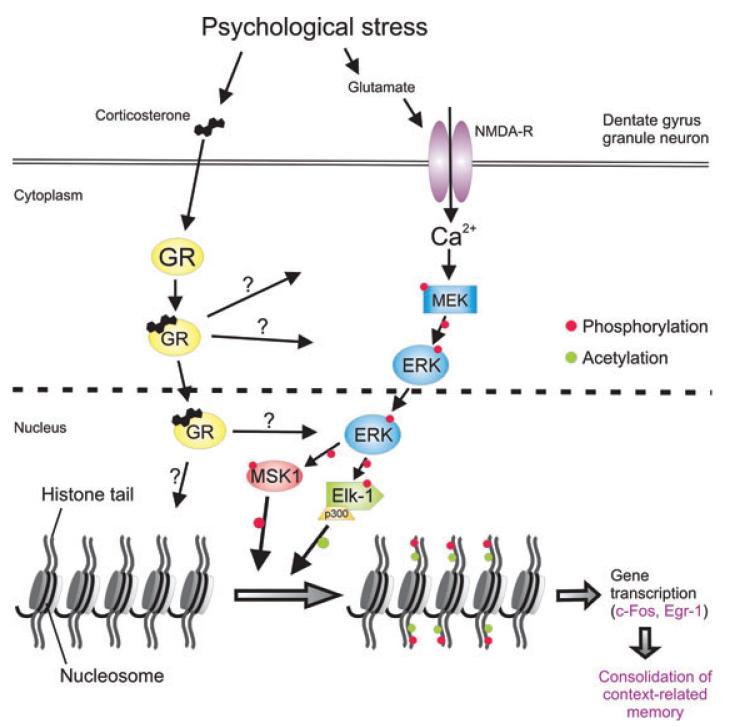
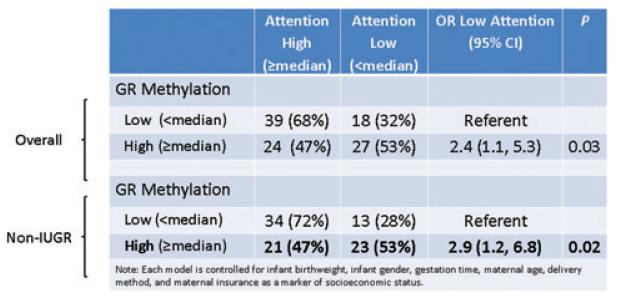
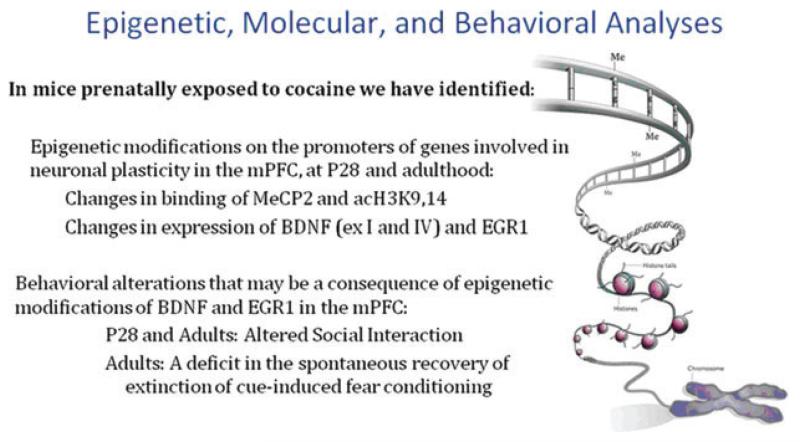
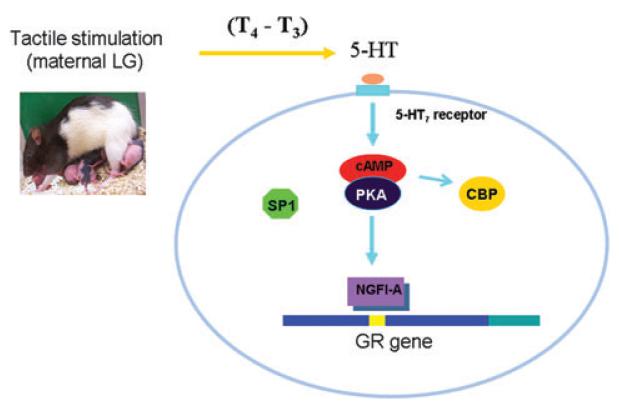
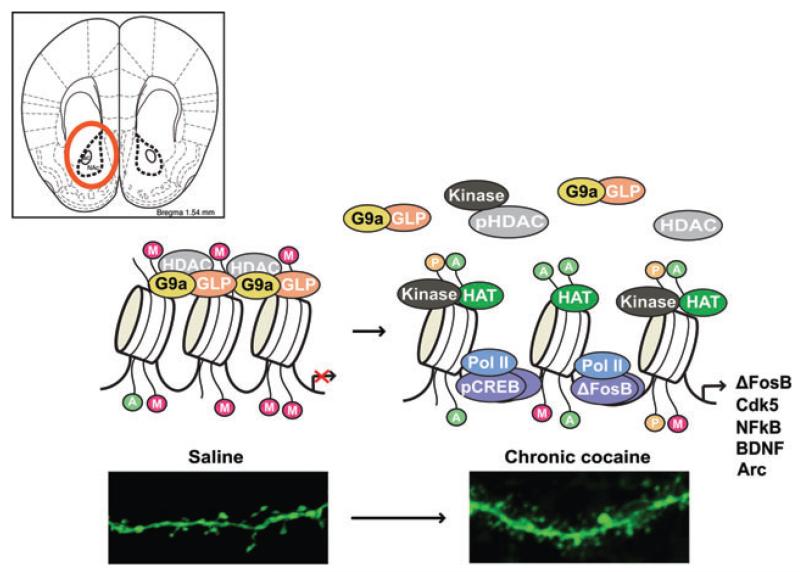
References
Appendix: ninety-six articles published to date on behavioral epigenetics
-
- Fischer A, Sananbenesi F, Wang X, et al. Recovery of learning and memory is associated with chromatin remodeling. Nature. 2007;447:178–182. - PubMed
-
- Tsankova NM, et al. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat. Neurosci. 2006;9:519–525. - PubMed
-
- Jakobsson J, et al. KAP1-mediated epigenetic repression in the forebrain modulates behavioral vulnerability to stress. Neuron. 2008;60:818–831. - PubMed
References
-
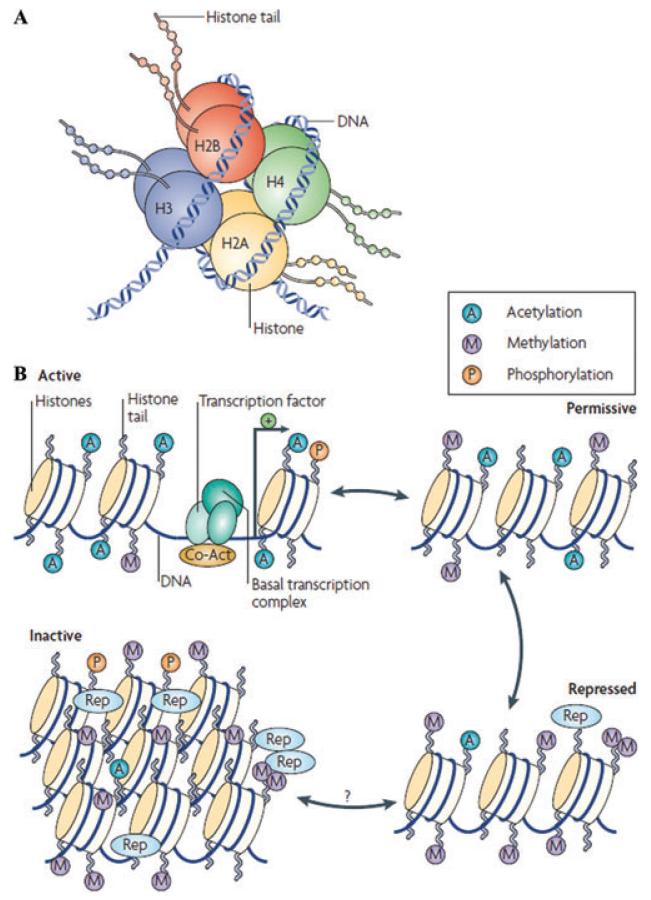
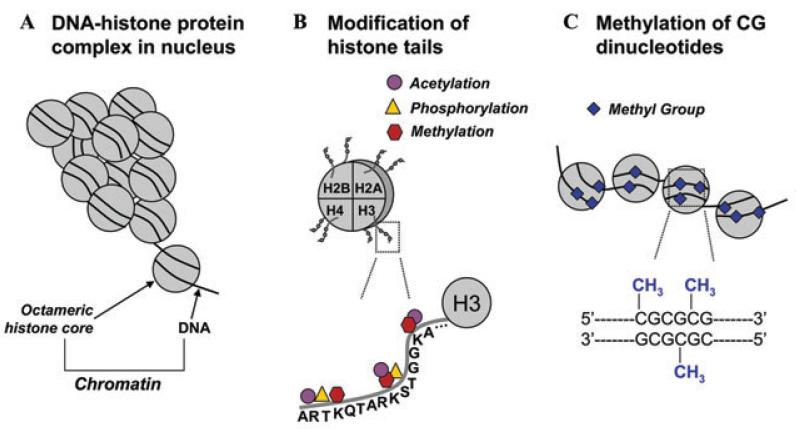
- Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. - PubMed
-
- Tsankova N, Renthal W, Kumar A, Nestler EJ. Epigenetic regulation in psychiatric disorders. Nature Rev. Neurosci. 2007;8:355–367. - PubMed
-
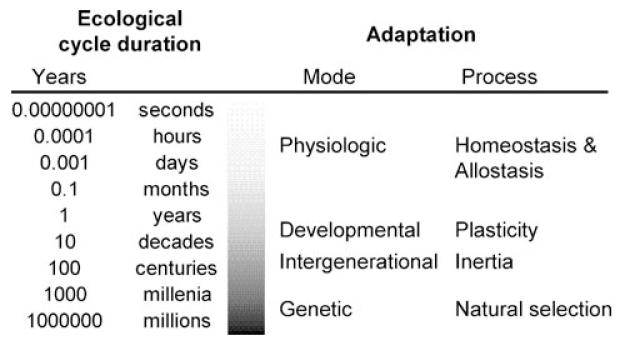
- Kuzawa CW, Quinn EA. Developmental origins of adult function and health: evolutionary hypotheses. Annu. Rev. Anthropol. 2009;38:131–147.
-
- Kuzawa CW. The fetal origins of developmental plasticity: are fetal cues reliable predictors of future nutritional environments? Am. J. Hum. Biol. 2005;17:5–21. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
