

Mechanisms of specificity in neuronal activity-regulated gene transcription
- PMID: 21620929
- PMCID: PMC3134613
- DOI: 10.1016/j.pneurobio.2011.05.003
Mechanisms of specificity in neuronal activity-regulated gene transcription
Abstract
The brain is a highly adaptable organ that is capable of converting sensory information into changes in neuronal function. This plasticity allows behavior to be accommodated to the environment, providing an important evolutionary advantage. Neurons convert environmental stimuli into long-lasting changes in their physiology in part through the synaptic activity-regulated transcription of new gene products. Since the neurotransmitter-dependent regulation of Fos transcription was first discovered nearly 25 years ago, a wealth of studies have enriched our understanding of the molecular pathways that mediate activity-regulated changes in gene transcription. These findings show that a broad range of signaling pathways and transcriptional regulators can be engaged by neuronal activity to sculpt complex programs of stimulus-regulated gene transcription. However, the shear scope of the transcriptional pathways engaged by neuronal activity raises the question of how specificity in the nature of the transcriptional response is achieved in order to encode physiologically relevant responses to divergent stimuli. Here we summarize the general paradigms by which neuronal activity regulates transcription while focusing on the molecular mechanisms that confer differential stimulus-, cell-type-, and developmental-specificity upon activity-regulated programs of neuronal gene transcription. In addition, we preview some of the new technologies that will advance our future understanding of the mechanisms and consequences of activity-regulated gene transcription in the brain.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
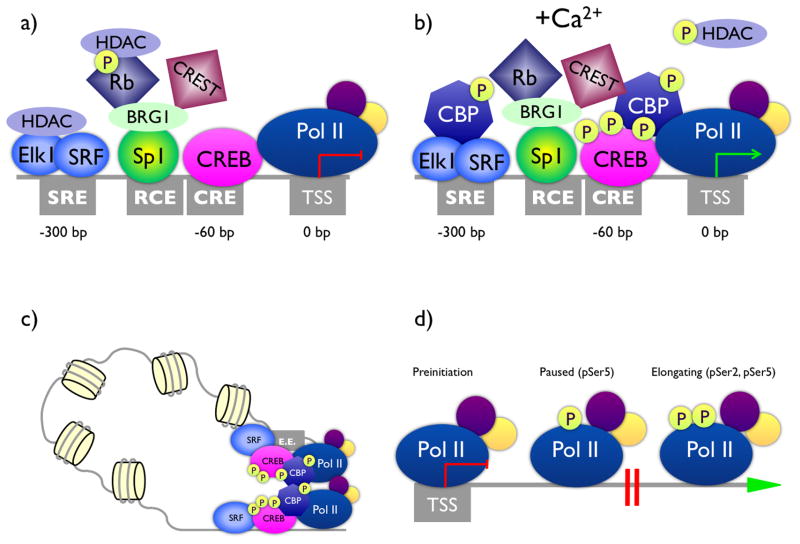
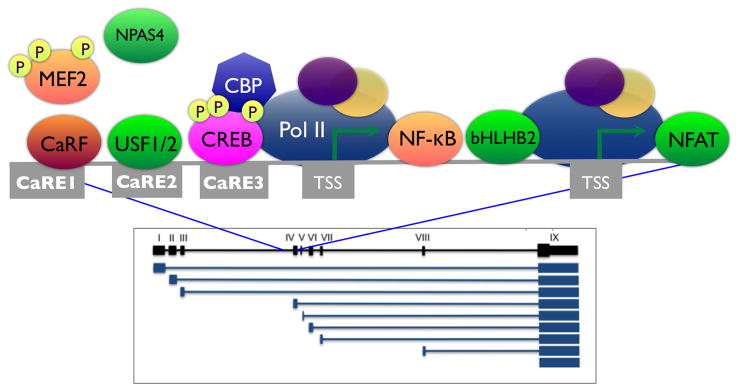
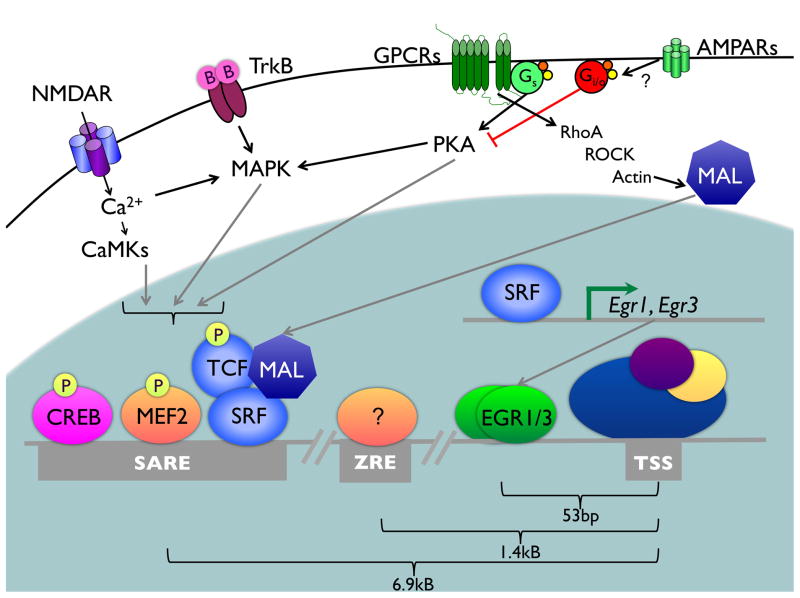
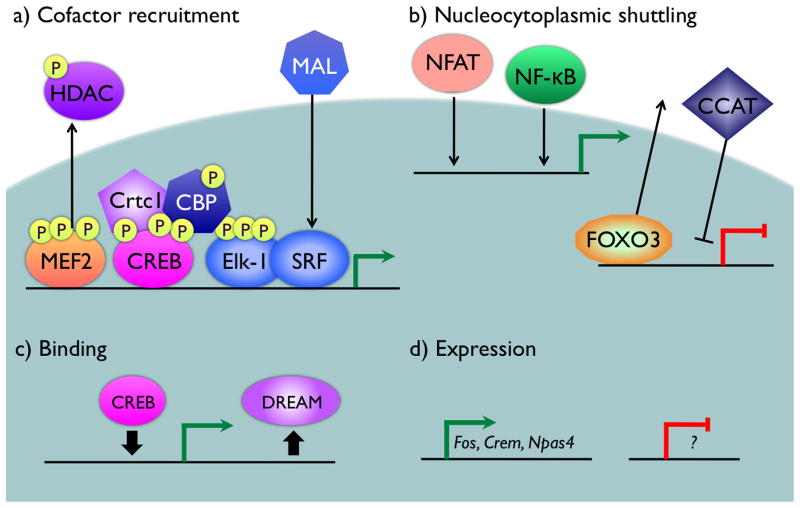
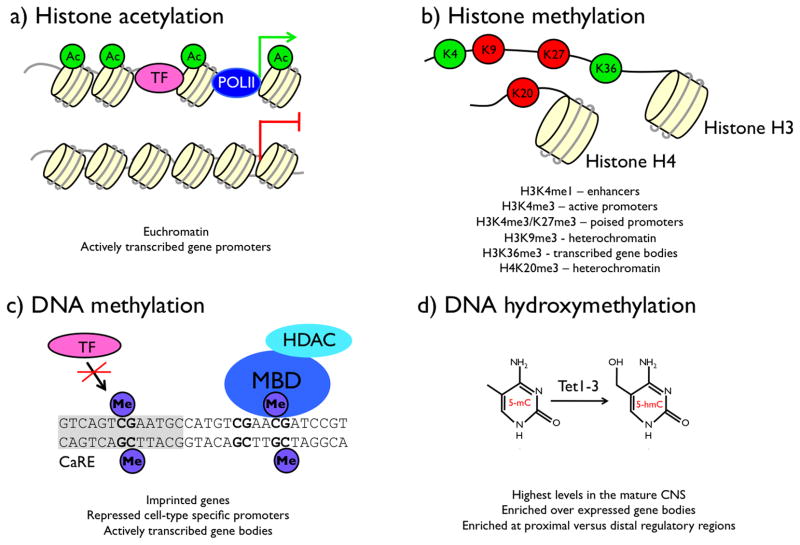
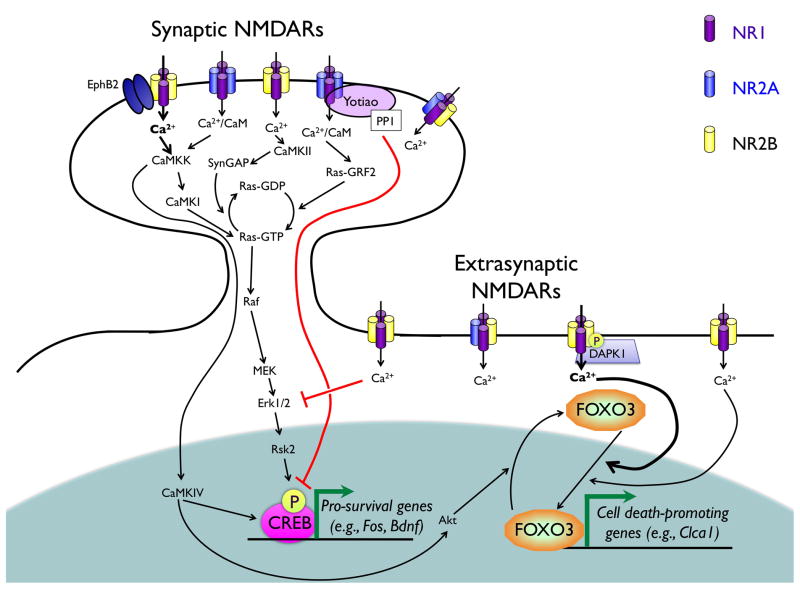
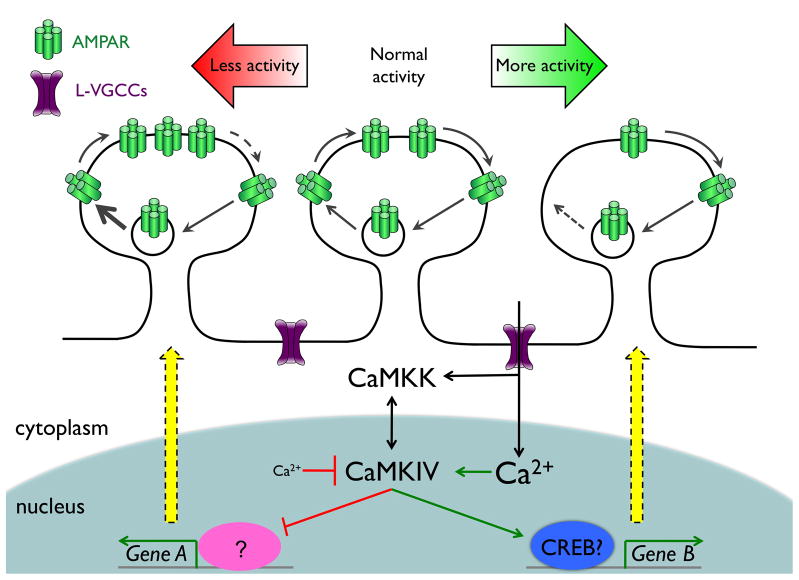
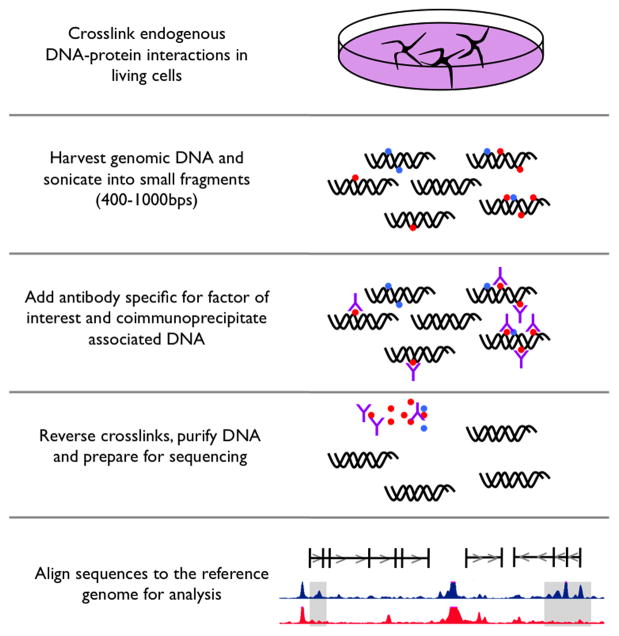
References
-
- Adams JP, Dudek SM. Late-phase long-term potentiation: getting to the nucleus. Nat Rev Neurosci. 2005;6:737–743. - PubMed
-
- Al-Hallaq RA, Jarabek BR, Fu Z, Vicini S, Wolfe BB, Yasuda RP. Association of NR3A with the N-methyl-D-aspartate receptor NR1 and NR2 subunits. Mol Pharmacol. 2002;62:1119–1127. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
