

Retroviral integrases promote fraying of viral DNA ends
- PMID: 21622554
- PMCID: PMC3138259
- DOI: 10.1074/jbc.M111.229179
Retroviral integrases promote fraying of viral DNA ends
Abstract
In the initial step of integration, retroviral integrase (IN) introduces precise nicks in the degenerate, short inverted repeats at the ends of linear viral DNA. The scissile phosphodiester bond is located immediately 3' of a highly conserved CA/GT dinucleotide, usually 2 bp from the ends. These nicks create new recessed 3'-OH viral DNA ends that are required for joining to host cell DNA. Previous studies have indicated that unpairing, "fraying," of the viral DNA ends by IN contributes to end recognition or catalysis. Here, we report that end fraying can be detected independently of catalysis with both avian sarcoma virus (ASV) and human immunodeficiency virus type 1 (HIV-1) IN proteins by use of fluorescence resonance energy transfer (FRET). The results were indicative of an IN-induced intramolecular conformational change in the viral DNA ends (cis FRET). Fraying activity is tightly coupled to the DNA binding capabilities of these enzymes, as follows: an inhibitor effective against both IN proteins was shown to block ASV IN DNA binding and end fraying, with similar dose responses; ASV IN substitutions that reduced DNA binding also reduced end fraying activity; and HIV-1 IN DNA binding and end fraying were both undetectable in the absence of a metal cofactor. Consistent with our previous results, end fraying is sequence-independent, suggesting that the DNA terminus per se is a major structural determinant for recognition. We conclude that frayed ends represent a functional intermediate in which DNA termini can be sampled for suitability for endonucleolytic processing.
Figures
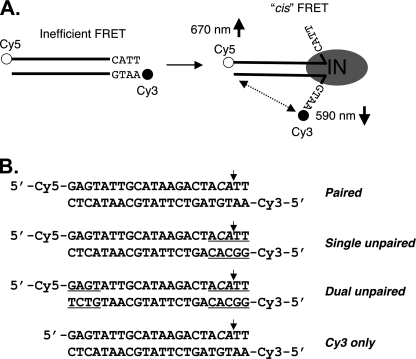
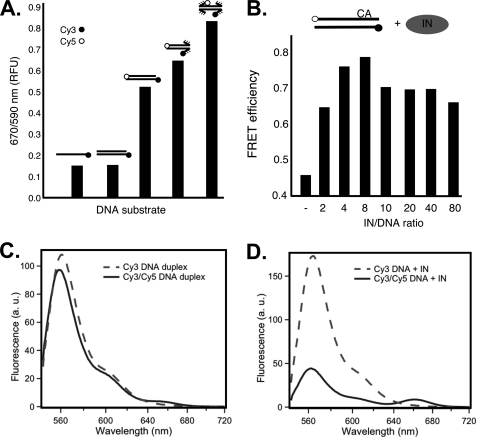
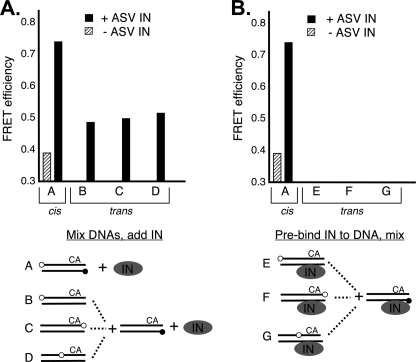
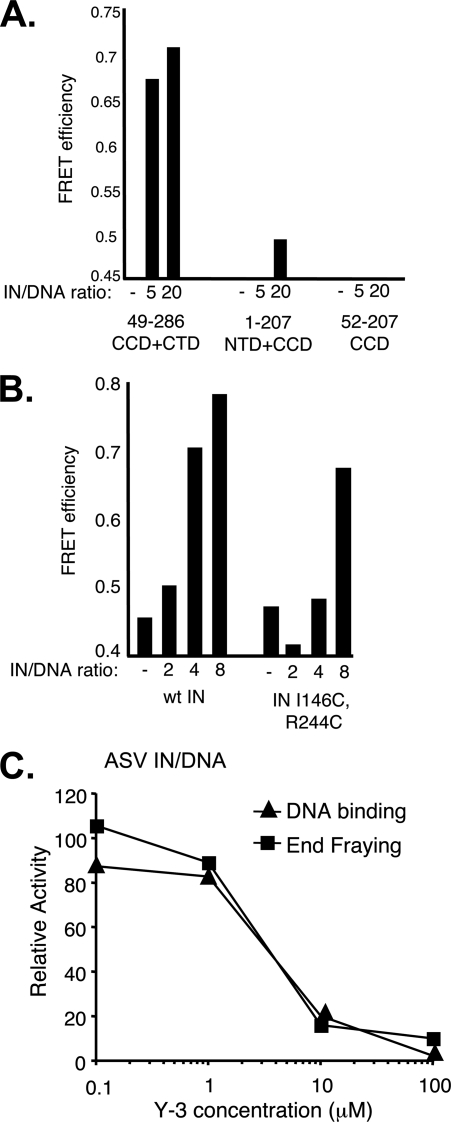
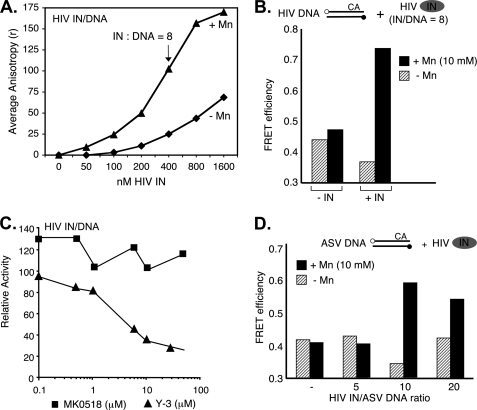
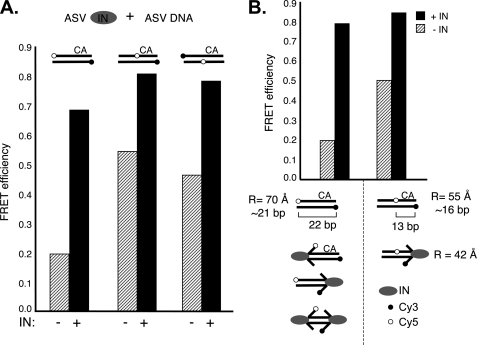
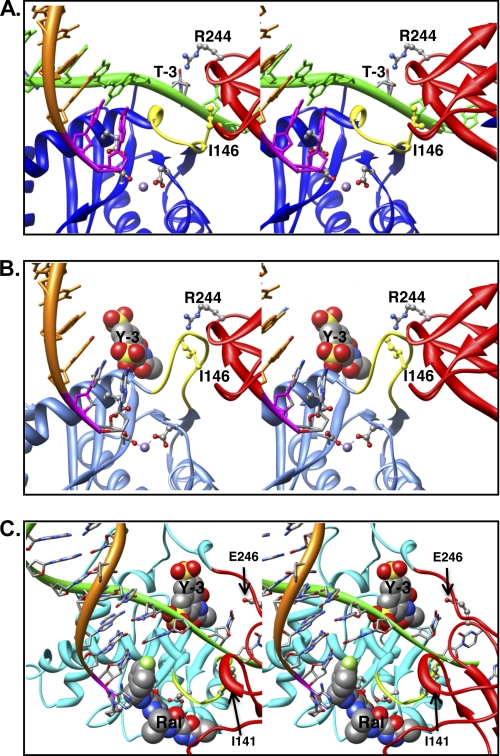
References
-
- Vandegraaff N., Engelman A. (2007) Expert Rev. Mol. Med. 9, 1–19 - PubMed
-
- Skalka A. M., Katz R. A. (2005) Cell Death Differ. 12, 971–978 - PubMed
-
- Yi J., Asante-Appiah E., Skalka A. M. (1999) Biochemistry 38, 8458–8468 - PubMed
-
- Katz R. A., DiCandeloro P., Kukolj G., Skalka A. M. (2001) J. Biol. Chem. 276, 34213–34220 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
