

Differential molecular responses of rice and wheat coleoptiles to anoxia reveal novel metabolic adaptations in amino acid metabolism for tissue tolerance
- PMID: 21622811
- PMCID: PMC3149938
- DOI: 10.1104/pp.111.175570
Differential molecular responses of rice and wheat coleoptiles to anoxia reveal novel metabolic adaptations in amino acid metabolism for tissue tolerance
Abstract
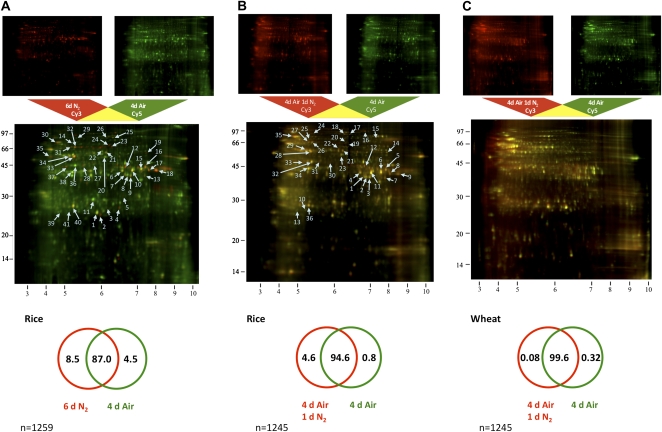
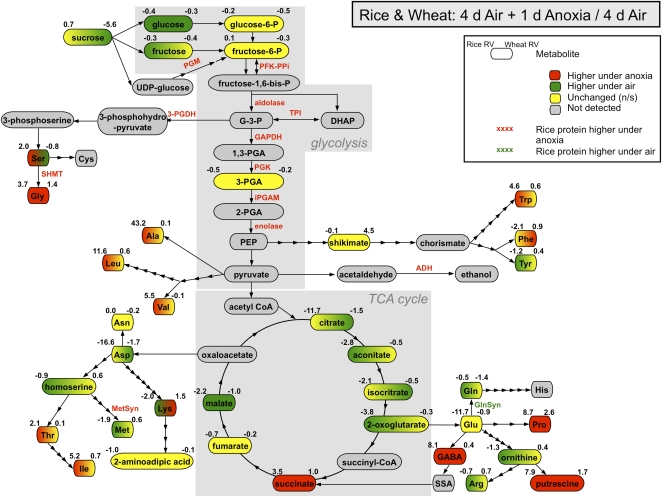
Rice (Oryza sativa) and wheat (Triticum aestivum) are the most important starch crops in world agriculture. While both germinate with an anatomically similar coleoptile, this tissue defines the early anoxia tolerance of rice and the anoxia intolerance of wheat seedlings. We combined protein and metabolite profiling analysis to compare the differences in response to anoxia between the rice and wheat coleoptiles. Rice coleoptiles responded to anoxia dramatically, not only at the level of protein synthesis but also at the level of altered metabolite pools, while the wheat response to anoxia was slight in comparison. We found significant increases in the abundance of proteins in rice coleoptiles related to protein translation and antioxidant defense and an accumulation of a set of enzymes involved in serine, glycine, and alanine biosynthesis from glyceraldehyde-3-phosphate or pyruvate, which correlates with an observed accumulation of these amino acids in anoxic rice. We show a positive effect on wheat root anoxia tolerance by exogenous addition of these amino acids, indicating that their synthesis could be linked to rice anoxia tolerance. The potential role of amino acid biosynthesis contributing to anoxia tolerance in cells is discussed.
Figures


References
-
- Atwell B, Waters I, Greenway H. (1982) The effect of oxygen and turbulence on elongation of coleoptiles of submergence-tolerant and -intolerant rice cultivars. J Exp Bot 33: 1030–1044
-
- Bailey-Serres J. (1999) Selective translation of cytoplasmic mRNAs in plants. Trends Plant Sci 4: 142–148 - PubMed
-
- Bailey-Serres J, Voesenek LACJ. (2008) Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 59: 313–339 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
