

Hierarchical organization and early hematopoietic specification of the developing HSC lineage in the AGM region
- PMID: 21624936
- PMCID: PMC3173253
- DOI: 10.1084/jem.20102419
Hierarchical organization and early hematopoietic specification of the developing HSC lineage in the AGM region
Abstract
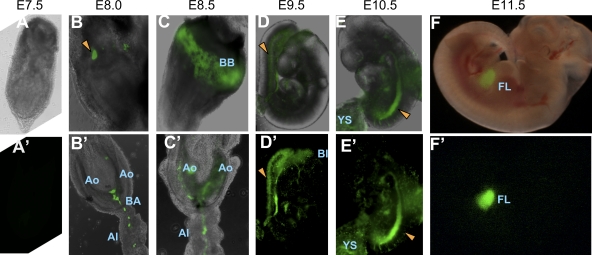
The aorta-gonad-mesonephros region plays an important role in hematopoietic stem cell (HSC) development during mouse embryogenesis. The vascular endothelial cadherin⁺ CD45⁺ (VE-cad⁺CD45⁺) population contains the major type of immature pre-HSCs capable of developing into long-term repopulating definitive HSCs. In this study, we developed a new coaggregation culture system, which supports maturation of a novel population of CD45-negative (VE-cad⁺CD45⁻CD41⁺) pre-HSCs into definitive HSCs. The appearance of these pre-HSCs precedes development of the VE-cad⁺CD45⁺ pre-HSCs (termed here type I and type II pre-HSCs, respectively), thus establishing a hierarchical directionality in the developing HSC lineage. By labeling the luminal surface of the dorsal aorta, we show that both type I and type II pre-HSCs are distributed broadly within the endothelial and subendothelial aortic layers, in contrast to mature definitive HSCs which localize to the aortic endothelial layer. In agreement with expression of CD41 in pre-HSCs, in vivo CD41-Cre-mediated genetic tagging occurs in embryonic pre-HSCs and persists in all lymphomyeloid lineages of the adult animal.
Figures







References
-
- Breviario F., Caveda L., Corada M., Martin-Padura I., Navarro P., Golay J., Introna M., Gulino D., Lampugnani M.G., Dejana E. 1995. Functional properties of human vascular endothelial cadherin (7B4/cadherin-5), an endothelium-specific cadherin. Arterioscler. Thromb. Vasc. Biol. 15:1229–1239 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
