

Pathogenity of some limb girdle muscular dystrophy mutations can result from reduced anchorage to myofibrils and altered stability of calpain 3
- PMID: 21624972
- PMCID: PMC3153300
- DOI: 10.1093/hmg/ddr239
Pathogenity of some limb girdle muscular dystrophy mutations can result from reduced anchorage to myofibrils and altered stability of calpain 3
Abstract
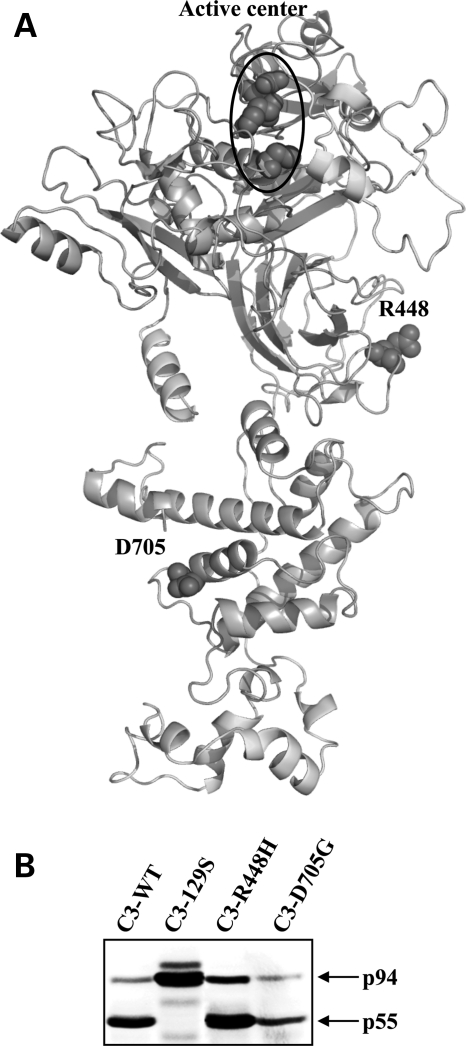
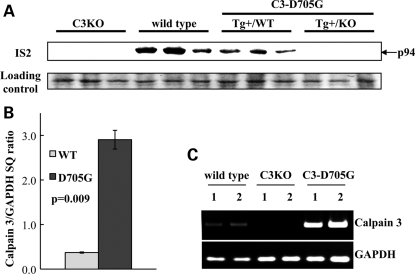
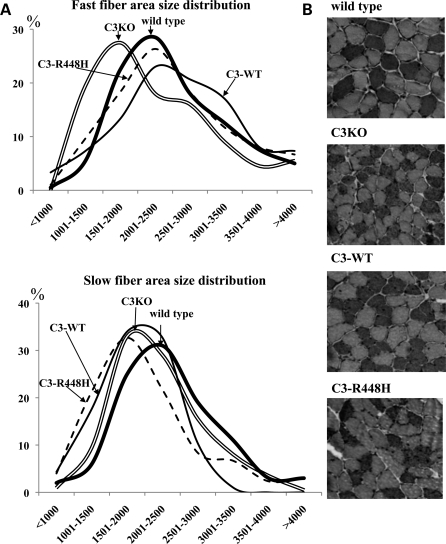
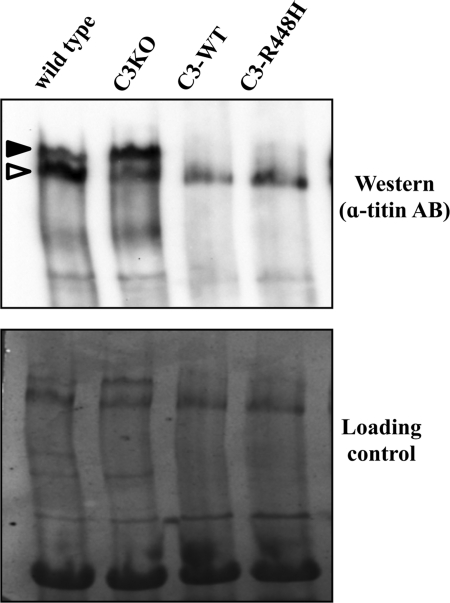
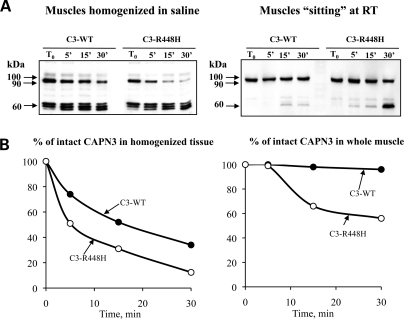
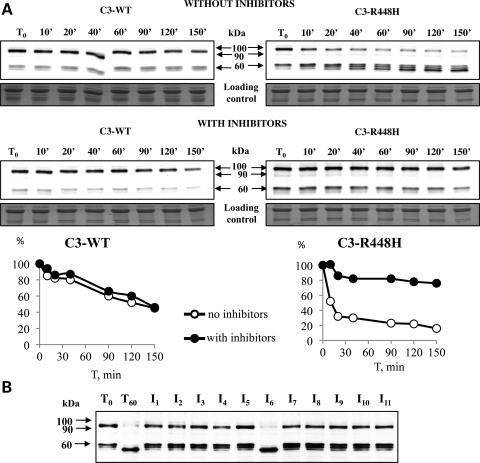
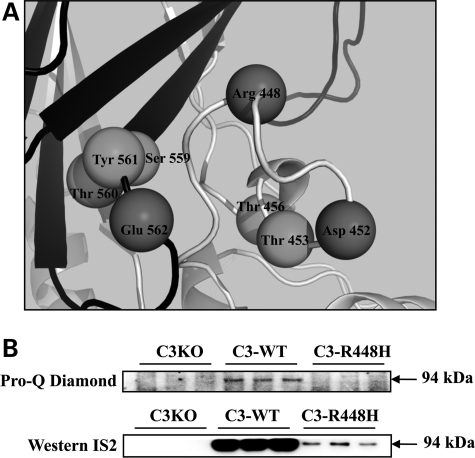
Calpain 3 (CAPN3) is a muscle-specific, calcium-dependent proteinase that is mutated in Limb Girdle Muscle Dystrophy type 2A. Most pathogenic missense mutations in LGMD2A affect CAPN3's proteolytic activity; however, two mutations, D705G and R448H, retain activity but nevertheless cause muscular dystrophy. Previously, we showed that D705G and R448H mutations reduce CAPN3s ability to bind to titin in vitro. In this investigation, we tested the consequence of loss of titin binding in vivo and examined whether this loss can be an underlying pathogenic mechanism in LGMD2A. To address this question, we created transgenic mice that express R448H or D705G in muscles, on wild-type (WT) CAPN3 or knock-out background. Both mutants were readily expressed in insect cells, but when D705G was expressed in skeletal muscle, it was not stable enough to study. Moreover, the D705G mutation had a dominant negative effect on endogenous CAPN3 when expressed on a WT background. The R448H protein was stably expressed in muscles; however, it was more rapidly degraded in muscle extracts compared with WT CAPN3. Increased degradation of R448H was due to non-cysteine, cellular proteases acting on the autolytic sites of CAPN3, rather than autolysis. Fractionation experiments revealed a significant decrease of R448H from the myofibrillar fraction, likely due to the mutant's inability to bind titin. Our data suggest that R448H and D705G mutations affect both CAPN3s anchorage to titin and its stability. These studies reveal a novel mechanism by which mutations that spare enzymatic activity can still lead to calpainopathy.
Figures



References
-
- Goll D.E., Thompson V.F., Li H., Wei W., Cong J. The calpain system. Physiol. Rev. 2003;83:731–801. - PubMed
-
- Sorimachi H., Hata S., Ono Y. Expanding members and roles of the calpain superfamily and their genetically modified animals. Exp. Anim. 2010;59:549–566. doi:10.1538/expanim.59.549. - DOI - PubMed
-
- Croall D.E., Ersfeld K. The calpains: modular designs and functional diversity. Genome Biol. 2007;8:218. doi:10.1186/gb-2007-8-6-218. - DOI - PMC - PubMed
-
- Sorimachi H., Suzuki K. The structure of calpain. J. Biochem. 2001;129:653–664. - PubMed
-
- Suzuki K., Sorimachi H., Yoshizawa T., Kinbara K., Ishiura S. Calpain: novel family members, activation, and physiologic function. Biol. Chem. Hoppe Seyler. 1995;376:523–529. doi:10.1515/bchm3.1995.376.9.523. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
