

Redistribution of caveolae during mitosis
- PMID: 21625007
- PMCID: PMC3104031
- DOI: 10.1242/jcs.076570
Redistribution of caveolae during mitosis
Abstract
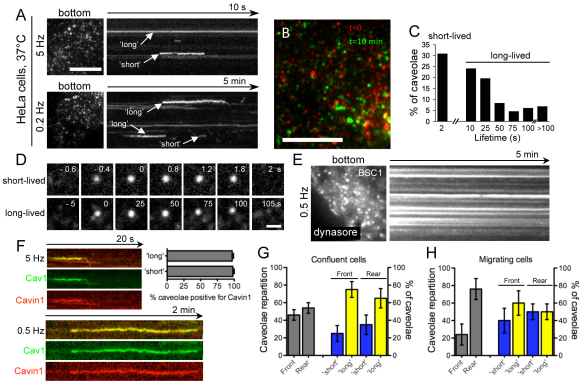
Caveolae form a specialized platform within the plasma membrane that is crucial for an array of important biological functions, ranging from signaling to endocytosis. Using total internal reflection fluorescence (TIRF) and 3D fast spinning-disk confocal imaging to follow caveola dynamics for extended periods, and electron microscopy to obtain high resolution snapshots, we found that the vast majority of caveolae are dynamic with lifetimes ranging from a few seconds to several minutes. Use of these methods revealed a change in the dynamics and localization of caveolae during mitosis. During interphase, the equilibrium between the arrival and departure of caveolae from the cell surface maintains the steady-state distribution of caveolin-1 (Cav1) at the plasma membrane. During mitosis, increased dynamics coupled to an imbalance between the arrival and departure of caveolae from the cell surface induces a redistribution of Cav1 from the plasma membrane to intracellular compartments. These changes are reversed during cytokinesis. The observed redistribution of Cav1 was reproduced by treatment of interphase cells with nocodazole, suggesting that microtubule rearrangements during mitosis can mediate caveolin relocalization. This study provides new insights into the dynamics of caveolae and highlights precise regulation of caveola budding and recycling during mitosis.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
