

Adam17-dependent shedding limits early neutrophil influx but does not alter early monocyte recruitment to inflammatory sites
- PMID: 21628404
- PMCID: PMC3142912
- DOI: 10.1182/blood-2010-11-321406
Adam17-dependent shedding limits early neutrophil influx but does not alter early monocyte recruitment to inflammatory sites
Abstract
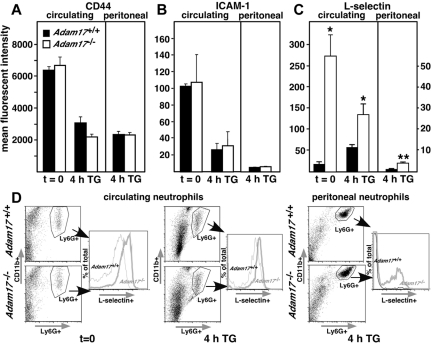
TNF-α-converting enzyme (TACE, herein denoted as Adam17) proteolytically sheds several cell-surface inflammatory proteins, but the physiologic importance of the cleavage of these substrates from leukocyte subsets during inflammation is incompletely understood. In this study, we show that Adam17-null neutrophils have a 2-fold advantage in their initial recruitment during thioglycollate-induced peritonitis, and they roll slower and adhere more readily in the cremaster model than wild-type neutrophils. Although CD44 and ICAM-1 are both in vitro substrates of Adam17, their surface levels are not altered on Adam17-null neutrophils. In contrast, L-selectin levels are elevated up to 10-fold in Adam17-null circulating neutrophils, and their accelerated peritoneal influx, slower rolling, and increased adhesion in the cremaster muscle are dependent on L-selectin. Analysis of mixed chimeras shows that enhanced L-selectin levels and accelerated influx were both cell-intrinsic properties of neutrophils lacking Adam17. In contrast, Adam17-null monocytes display no acceleration of infiltration into the peritoneum in spite of elevated L-selectin surface levels, and their peritoneal influx was independent of L-selectin. Therefore, our data demonstrate substrate and myeloid cell-type specificity of Adam17-mediated cleavage of its substrates, and show that neutrophils and monocytes use distinct mechanisms for infiltration of tissues.
Figures





References
-
- Butcher EC. Leukocyte-endothelial cell recognition: three (or more) steps to specificity and diversity. Cell. 1991;67(6):1033–1036. - PubMed
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76(2):301–314. - PubMed
-
- Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7(9):678–689. - PubMed
-
- Dunon D, Piali L, Imhof BA. To stick or not to stick: the new leukocyte homing paradigm. Curr Opin Cell Biol. 1996;8(5):714–723. - PubMed
-
- Kishimoto TK, Jutila MA, Berg EL, Butcher EC. Neutrophil Mac-1 and MEL-14 adhesion proteins inversely regulated by chemotactic factors. Science. 1989;245(4923):1238–1241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
