

Pervasive haplotypic variation in the spliceo-transcriptome of the human major histocompatibility complex
- PMID: 21628452
- PMCID: PMC3129247
- DOI: 10.1101/gr.116681.110
Pervasive haplotypic variation in the spliceo-transcriptome of the human major histocompatibility complex
Abstract
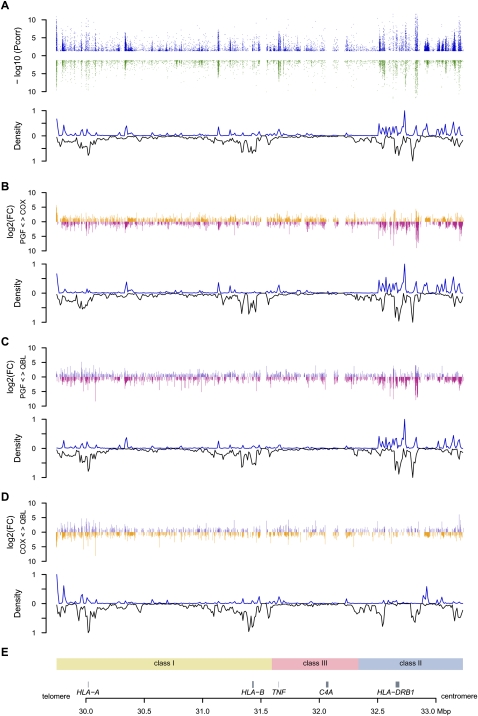
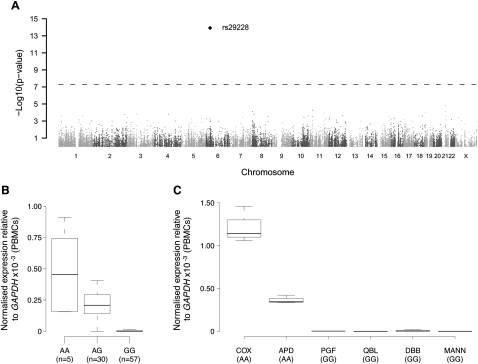
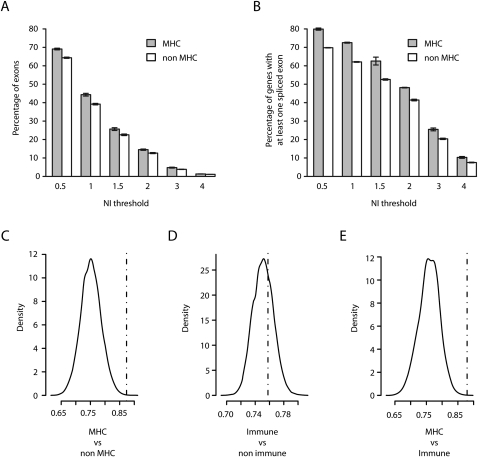
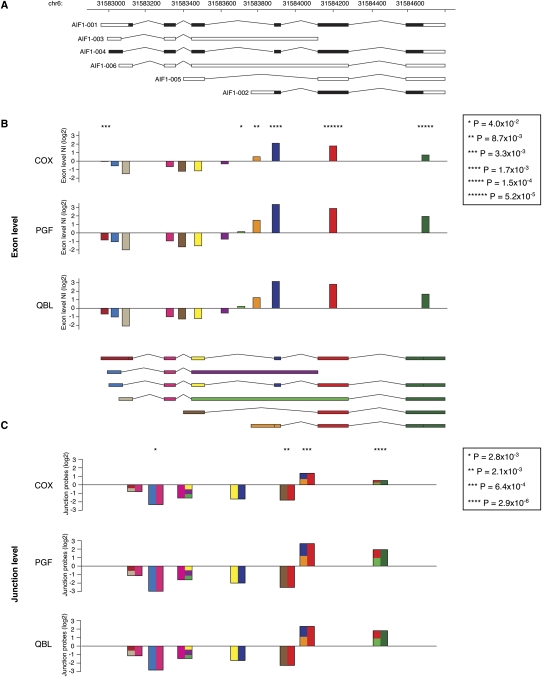
The human major histocompatibility complex (MHC) on chromosome 6p21 is a paradigm for genomics, showing remarkable polymorphism and striking association with immune and non-immune diseases. The complex genomic landscape of the MHC, notably strong linkage disequilibrium, has made resolving causal variants very challenging. A promising approach is to investigate gene expression levels considered as tractable intermediate phenotypes in mapping complex diseases. However, how transcription varies across the MHC, notably relative to specific haplotypes, remains unknown. Here, using an original hybrid tiling and splice junction microarray that includes alternate allele probes, we draw the first high-resolution strand-specific transcription map for three common MHC haplotypes (HLA-A1-B8-Cw7-DR3, HLA-A3-B7-Cw7-DR15, and HLA-A26-B18-Cw5-DR3-DQ2) strongly associated with autoimmune diseases including type 1 diabetes, systemic lupus erythematosus, and multiple sclerosis. We find that haplotype-specific differences in gene expression are common across the MHC, affecting 96 genes (46.4%), most significantly the zing finger protein gene ZFP57. Differentially expressed probes are correlated with polymorphisms between haplotypes, consistent with cis effects that we directly demonstrate for ZFP57 in a cohort of healthy volunteers (P = 1.2 × 10(-14)). We establish that alternative splicing is significantly more frequent in the MHC than genome-wide (72.5% vs. 62.1% of genes, P ≤ 1 × 10(-4)) and shows marked haplotypic differences. We also unmask novel and abundant intergenic transcription involving 31% of transcribed blocks identified. Our study reveals that the renowned MHC polymorphism also manifests as transcript diversity, and our novel haplotype-based approach marks a new step toward identification of regulatory variants involved in the control of MHC-associated phenotypes and diseases.
Figures

Comment in
-
Complex disease: ups and downs at the MHC.Nat Rev Genet. 2011 Jun 17;12(7):456-7. doi: 10.1038/nrg3021. Nat Rev Genet. 2011. PMID: 21681207 No abstract available.
References
-
- Ahmad T, Neville M, Marshall SE, Armuzzi A, Mulcahy-Hawes K, Crawshaw J, Sato H, Ling KL, Barnardo M, Goldthorpe S, et al. 2003. Haplotype-specific linkage disequilibrium patterns define the genetic topography of the human MHC. Hum Mol Genet 12: 647–656 - PubMed
-
- Bertone P, Stolc V, Royce TE, Rozowsky JS, Urban AE, Zhu X, Rinn JL, Tongprasit W, Samanta M, Weissman S, et al. 2004. Global identification of human transcribed sequences with genome tiling arrays. Science 306: 2242–2246 - PubMed
-
- Carosella ED, Favier B, Rouas-Freiss N, Moreau P, Lemaoult J 2008. Beyond the increasing complexity of the immunomodulatory HLA-G molecule. Blood 111: 4862–4870 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials