

Surface export of GAPDH/SDH, a glycolytic enzyme, is essential for Streptococcus pyogenes virulence
- PMID: 21628503
- PMCID: PMC3104492
- DOI: 10.1128/mBio.00068-11
Surface export of GAPDH/SDH, a glycolytic enzyme, is essential for Streptococcus pyogenes virulence
Abstract
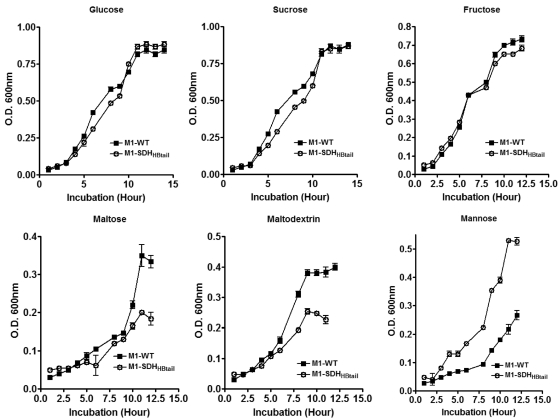
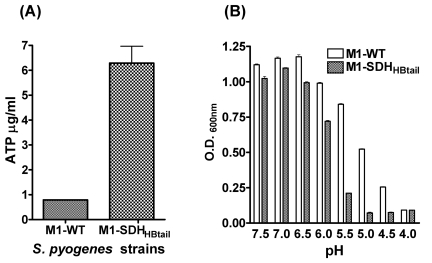
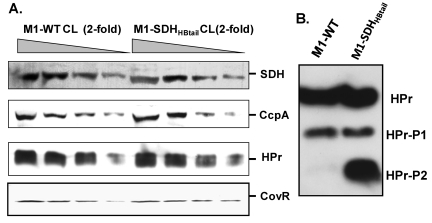
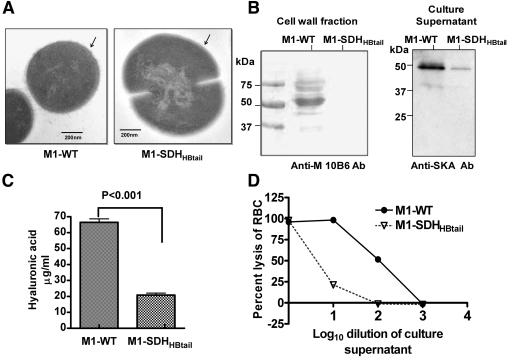
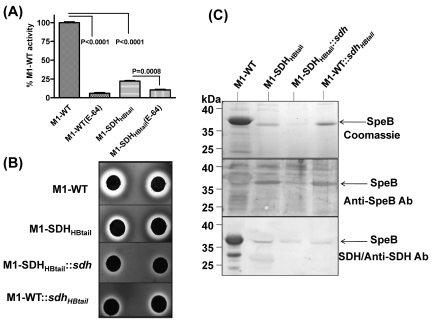
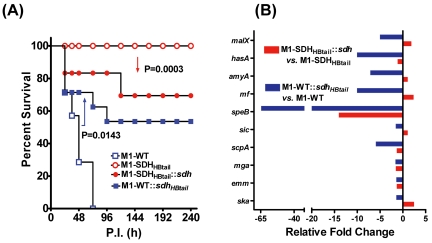
Streptococcal surface dehydrogenase (SDH) (glyceraldehyde-3-phosphate dehydrogenase [GAPDH]) is an anchorless major multifunctional surface protein in group A Streptococcus (GAS) with the ability to bind important mammalian proteins, including plasmin(ogen). Although several biological properties of SDH are suggestive of its possible role in GAS virulence, its direct role in GAS pathogenesis has not been ascertained because it is essential for GAS survival. Thus, it has remained enigmatic as to "how and why" SDH/GAPDH is exported onto the bacterial surface. The present investigation highlights "why" SDH is exported onto the GAS surface. Differential microarray-based genome-wide transcript abundance analysis was carried out using a specific mutant, which was created by inserting a hydrophobic tail at the C-terminal end of SDH (M1-SDH(HBtail)) and thus preventing its exportation onto the GAS surface. This analysis revealed downregulation of the majority of genes involved in GAS virulence and genes belonging to carbohydrate and amino acid metabolism and upregulation of those related to lipid metabolism. The complete attenuation of this mutant for virulence in the mouse model and the decreased and increased virulence of the wild-type and mutant strains postcomplementation with SDH(HBtail) and SDH, respectively, indicated that the SDH surface export indeed regulates GAS virulence. M1-SDH(HBtail) also displayed unaltered growth patterns, increased intracellular ATP concentration and Hpr double phosphorylation, and significantly reduced pH tolerance, streptolysin S, and SpeB activities. These phenotypic and physiological changes observed in the mutant despite the unaltered expression levels of established transcriptional regulators further highlight the fact that SDH interfaces with many regulators and its surface exportation is essential for GAS virulence.
Importance: Streptococcal surface dehydrogenase (SDH), a classical anchorless cytoplasmically localized glycolytic enzyme, is exported onto the group A Streptococcus (GAS) surface through a hitherto unknown mechanism(s). It has not been known why GAS or other prokaryotes should export this protein onto the surface. By genetic manipulations, we created a novel GAS mutant strain expressing SDH with a 12-amino-acid hydrophobic tail at its C-terminal end and thus were able to prevent its surface exportation without altering its enzymatic activity or growth pattern. Interestingly, the mutant was completely attenuated for virulence in a mouse peritonitis model. The global gene expression profiles of this mutant reveal that the surface exportation of SDH is mandatory to maintain GAS virulence. The ability of GAS as a successful pathogen to localize SDH in the cytoplasm as well as on the surface is physiologically relevant and dynamically obligatory to fine-tune the functions of many transcriptional regulators and also to exploit its virulence properties for infection.
Figures
Similar articles
-
Inhibition of cell surface export of group A streptococcal anchorless surface dehydrogenase affects bacterial adherence and antiphagocytic properties.Infect Immun. 2005 Oct;73(10):6237-48. doi: 10.1128/IAI.73.10.6237-6248.2005. Infect Immun. 2005. PMID: 16177295 Free PMC article.
-
Endopeptidase PepO Regulates the SpeB Cysteine Protease and Is Essential for the Virulence of Invasive M1T1 Streptococcus pyogenes.J Bacteriol. 2018 Mar 26;200(8):e00654-17. doi: 10.1128/JB.00654-17. Print 2018 Apr 15. J Bacteriol. 2018. PMID: 29378883 Free PMC article.
-
Multifunctional glyceraldehyde-3-phosphate dehydrogenase of Streptococcus pyogenes is essential for evasion from neutrophils.J Biol Chem. 2006 May 19;281(20):14215-23. doi: 10.1074/jbc.M513408200. Epub 2006 Mar 24. J Biol Chem. 2006. PMID: 16565520
-
Role of extracellular GAPDH in Streptococcus pyogenes virulence.Mo Med. 2013 May-Jun;110(3):236-40. Mo Med. 2013. PMID: 23829111 Free PMC article. Review.
-
Molecular insight into invasive group A streptococcal disease.Nat Rev Microbiol. 2011 Sep 16;9(10):724-36. doi: 10.1038/nrmicro2648. Nat Rev Microbiol. 2011. PMID: 21921933 Review.
Cited by
-
Streptococcus pneumoniae GAPDH Is Released by Cell Lysis and Interacts with Peptidoglycan.PLoS One. 2015 Apr 30;10(4):e0125377. doi: 10.1371/journal.pone.0125377. eCollection 2015. PLoS One. 2015. PMID: 25927608 Free PMC article.
-
Role of serine/threonine phosphatase (SP-STP) in Streptococcus pyogenes physiology and virulence.J Biol Chem. 2011 Dec 2;286(48):41368-41380. doi: 10.1074/jbc.M111.286690. Epub 2011 Sep 14. J Biol Chem. 2011. PMID: 21917918 Free PMC article.
-
Identification of GAPDH on the surface of Plasmodium sporozoites as a new candidate for targeting malaria liver invasion.J Exp Med. 2016 Sep 19;213(10):2099-112. doi: 10.1084/jem.20160059. Epub 2016 Aug 22. J Exp Med. 2016. PMID: 27551151 Free PMC article.
-
Gymnemic Acids Inhibit Adhesive Nanofibrillar Mediated Streptococcus gordonii-Candida albicans Mono-Species and Dual-Species Biofilms.Front Microbiol. 2019 Oct 11;10:2328. doi: 10.3389/fmicb.2019.02328. eCollection 2019. Front Microbiol. 2019. PMID: 31681200 Free PMC article.
-
Schistosoma mansoni glyceraldehyde-3-phosphate dehydrogenase enhances formation of the blood-clot lysis protein plasmin.Biol Open. 2020 Mar 24;9(3):bio050385. doi: 10.1242/bio.050385. Biol Open. 2020. PMID: 32098782 Free PMC article.
References
-
- Tart AH, Walker MJ, Musser JM. 2007. New understanding of the group A Streptococcus pathogenesis cycle. Trends Microbiol. 15:318–325 - PubMed
-
- Carapetis JR, Steer AC, Mulholland EK, Weber M. 2005. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 5:685–694 - PubMed
-
- Kreikemeyer B, McIver KS, Podbielski A. 2003. Virulence factor regulation and regulatory networks in Streptococcus pyogenes and their impact on pathogen-host interactions. Trends Microbiol. 11:224–232 - PubMed
-
- Churchward G. 2007. The two faces of Janus: virulence gene regulation by CovR/S in group A streptococci. Mol. Microbiol. 64:34–41 - PubMed
-
- McIver KS. 2009. Stand-alone response regulators controlling global virulence networks in Streptococcus pyogenes. Contrib. Microbiol. 16:103–119 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
