

Root cortical aerenchyma enhances the growth of maize on soils with suboptimal availability of nitrogen, phosphorus, and potassium
- PMID: 21628631
- PMCID: PMC3135917
- DOI: 10.1104/pp.111.175489
Root cortical aerenchyma enhances the growth of maize on soils with suboptimal availability of nitrogen, phosphorus, and potassium
Abstract
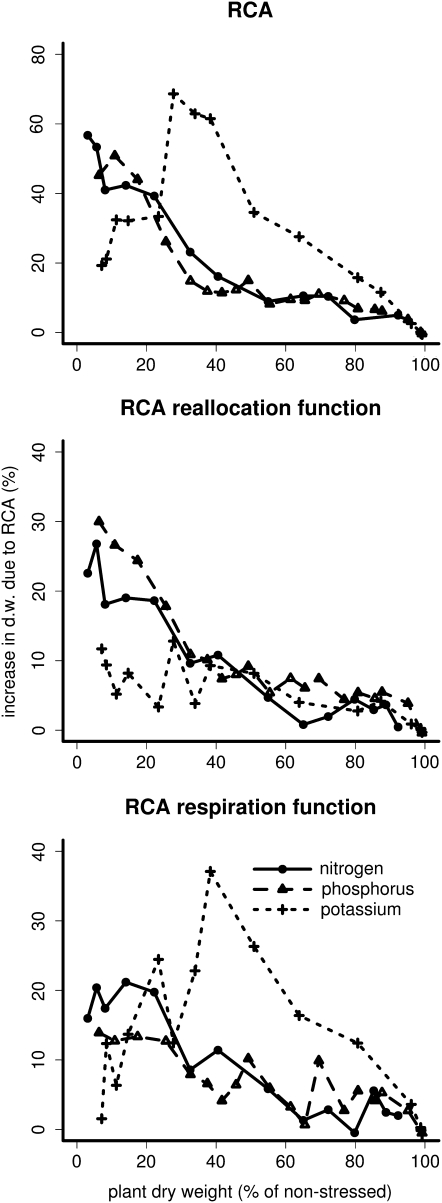
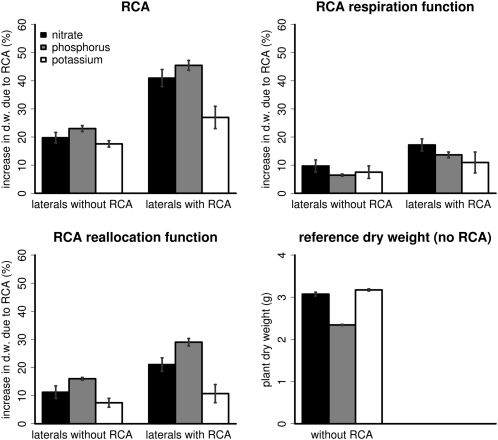
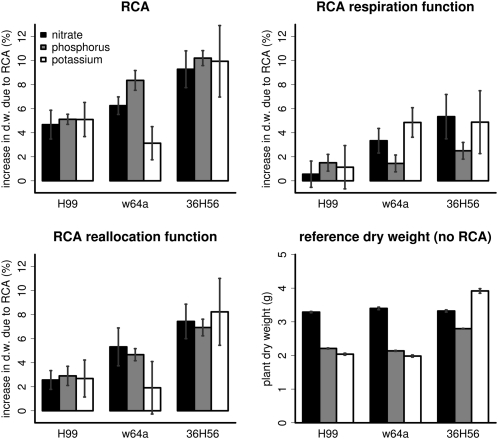
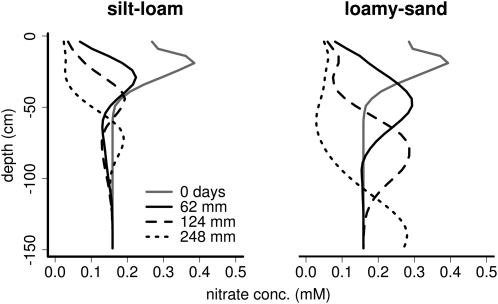
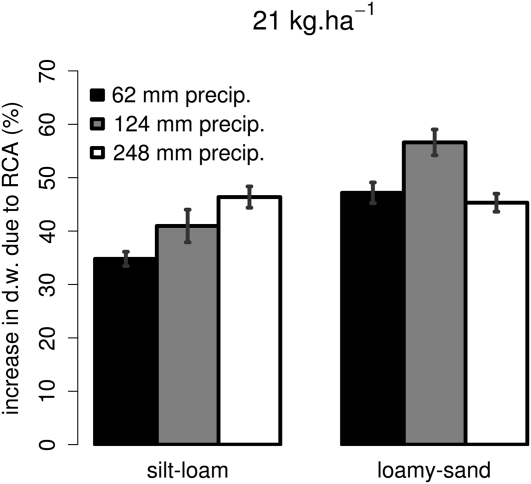
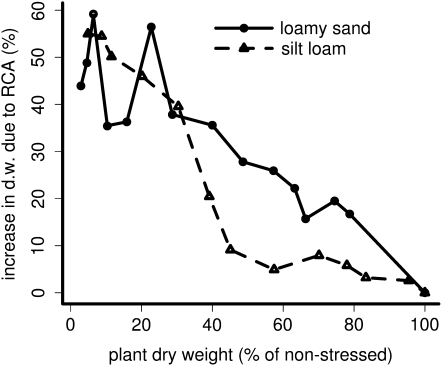
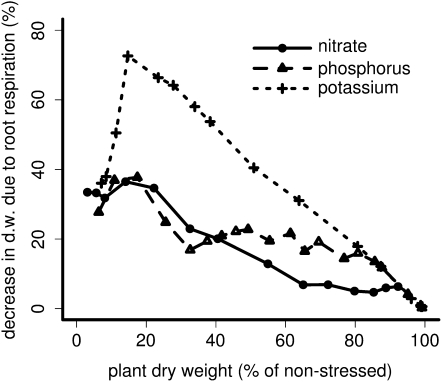
Root cortical aerenchyma (RCA) is induced by hypoxia, drought, and several nutrient deficiencies. Previous research showed that RCA formation reduces the respiration and nutrient content of root tissue. We used SimRoot, a functional-structural model, to provide quantitative support for the hypothesis that RCA formation is a useful adaptation to suboptimal availability of phosphorus, nitrogen, and potassium by reducing the metabolic costs of soil exploration in maize (Zea mays). RCA increased the growth of simulated 40-d-old maize plants up to 55%, 54%, or 72% on low nitrogen, phosphorus, or potassium soil, respectively, and reduced critical fertility levels by 13%, 12%, or 7%, respectively. The greater utility of RCA on low-potassium soils is associated with the fact that root growth in potassium-deficient plants was more carbon limited than in phosphorus- and nitrogen-deficient plants. In contrast to potassium-deficient plants, phosphorus- and nitrogen-deficient plants allocate more carbon to the root system as the deficiency develops. The utility of RCA also depended on other root phenes and environmental factors. On low-phosphorus soils (7.5 μM), the utility of RCA was 2.9 times greater in plants with increased lateral branching density than in plants with normal branching. On low-nitrate soils, the utility of RCA formation was 56% greater in coarser soils with high nitrate leaching. Large genetic variation in RCA formation and the utility of RCA for a range of stresses position RCA as an interesting crop-breeding target for enhanced soil resource acquisition.
Figures



References
-
- Borch K, Bouma TJ, Lynch JP, Brown KM. (1999) Ethylene: a regulator of root architectural responses to soil phosphorus availability. Plant Cell Environ 22: 425–431
-
- Bouranis DL, Chorianopoulou SN, Siyiannis VF, Protonotarios VE, Hawkesford MJ. (2003) Aerenchyma formation in roots of maize during sulphate starvation. Planta 217: 382–391 - PubMed
-
- Brown KM, Zhang YJ, Kim HJ, Lynch JP. (2003) The ethylene underground. Acta Hortic 618: 193–198
-
- Burton AL. (2010) Phenotypic evaluation and genetic basis of anatomical and architectural root traits in the genus Zea. PhD thesis. The Pennsylvania State University, University Park, PA
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
