

Ever-young sex chromosomes in European tree frogs
- PMID: 21629756
- PMCID: PMC3100596
- DOI: 10.1371/journal.pbio.1001062
Ever-young sex chromosomes in European tree frogs
Abstract
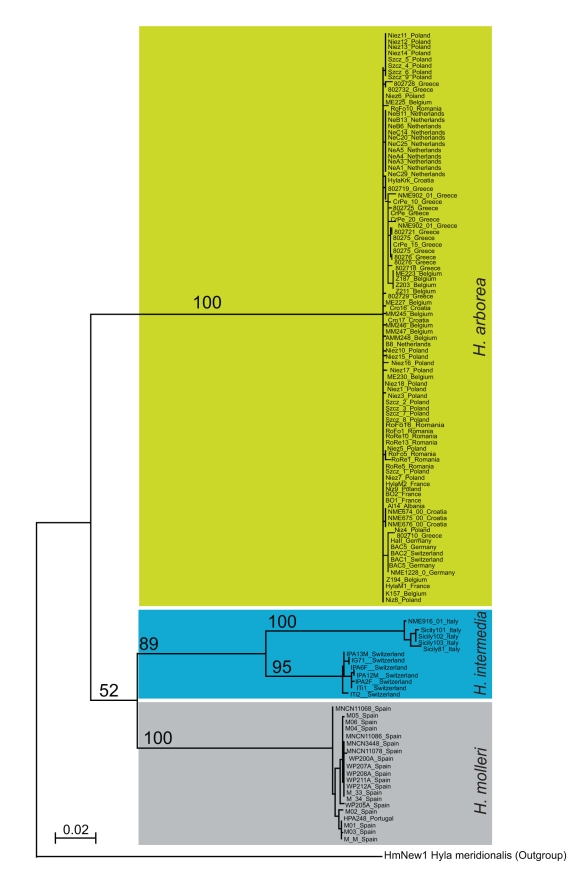
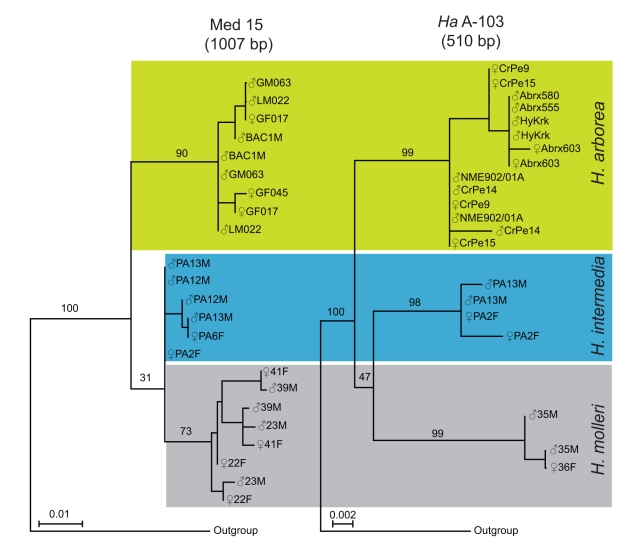
Non-recombining sex chromosomes are expected to undergo evolutionary decay, ending up genetically degenerated, as has happened in birds and mammals. Why are then sex chromosomes so often homomorphic in cold-blooded vertebrates? One possible explanation is a high rate of turnover events, replacing master sex-determining genes by new ones on other chromosomes. An alternative is that X-Y similarity is maintained by occasional recombination events, occurring in sex-reversed XY females. Based on mitochondrial and nuclear gene sequences, we estimated the divergence times between European tree frogs (Hyla arborea, H. intermedia, and H. molleri) to the upper Miocene, about 5.4-7.1 million years ago. Sibship analyses of microsatellite polymorphisms revealed that all three species have the same pair of sex chromosomes, with complete absence of X-Y recombination in males. Despite this, sequences of sex-linked loci show no divergence between the X and Y chromosomes. In the phylogeny, the X and Y alleles cluster according to species, not in groups of gametologs. We conclude that sex-chromosome homomorphy in these tree frogs does not result from a recent turnover but is maintained over evolutionary timescales by occasional X-Y recombination. Seemingly young sex chromosomes may thus carry old-established sex-determining genes, a result at odds with the view that sex chromosomes necessarily decay until they are replaced. This raises intriguing perspectives regarding the evolutionary dynamics of sexually antagonistic genes and the mechanisms that control X-Y recombination.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
-
- Lahn B. T, Page D. C. Four evolutionary strata on the human X chromosome. Science. 1999;286:964–967. - PubMed
-
- Graves J. A. M. Sex chromosome specialization and degeneration in mammals. Cell. 2006;124:901–914. - PubMed
-
- Rice W. R. Evolution of the Y sex chromosome in animals. Bioscience. 1996;46:331–343.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
