

The earliest intrathymic precursors of CD8α(+) thymic dendritic cells correspond to myeloid-type double-negative 1c cells
- PMID: 21630253
- PMCID: PMC4291128
- DOI: 10.1002/eji.201141728
The earliest intrathymic precursors of CD8α(+) thymic dendritic cells correspond to myeloid-type double-negative 1c cells
Abstract
The dendritic cells (DCs) present in lymphoid and non-lymphoid organs are generated from progenitors with myeloid-restricted potential. However, in the thymus a major subset of DCs expressing CD8α and langerin (CD207) appears to stand apart from all other DCs in that it is thought to derive from progenitors with lymphoid potential. Using mice expressing a fluorescent reporter and a diphtheria toxin receptor under the control of the cd207 gene, we demonstrated that CD207(+) CD8α(+) thymic DCs do not share a common origin with T cells but originate from intrathymic precursors that express markers that are normally present on all (CD11c(+) and MHCII molecules) or on some (CD207, CD135, CD8α, CX3CR1) DC subsets. Those intrathymic myeloid-type precursors correspond to CD44(+) CD25(-) double-negative 1c (DN1c) cells and are continuously renewed from bone marrow-derived canonical DC precursors. In conclusion, our results demonstrate that the earliest intrathymic precursors of CD8α(+) thymic DCs correspond to myeloid-type DN1c cells and support the view that under physiological conditions myeloid-restricted progenitors generate the whole constellation of DCs present in the body including the thymus.
Copyright © 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Conflict of interest statement
Conflict of interest: The authors declare no financial or commercial conflict of interest.
Figures





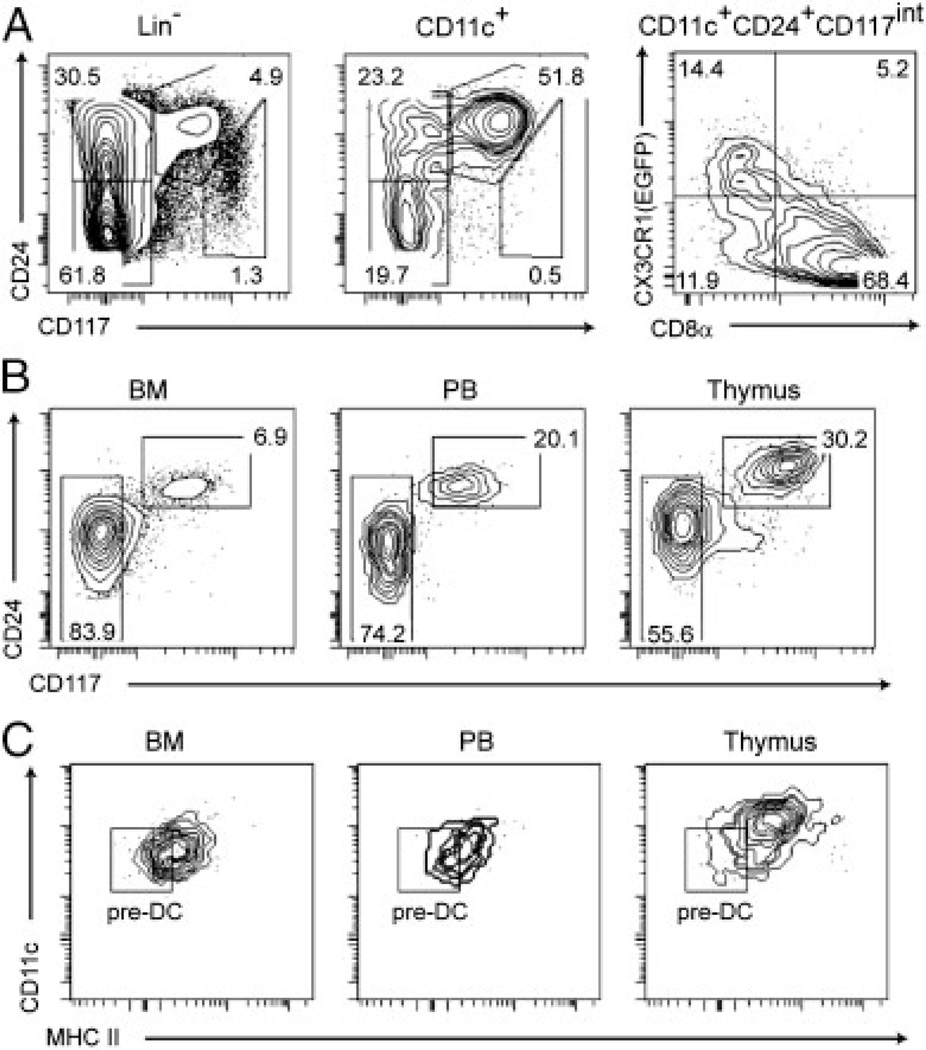

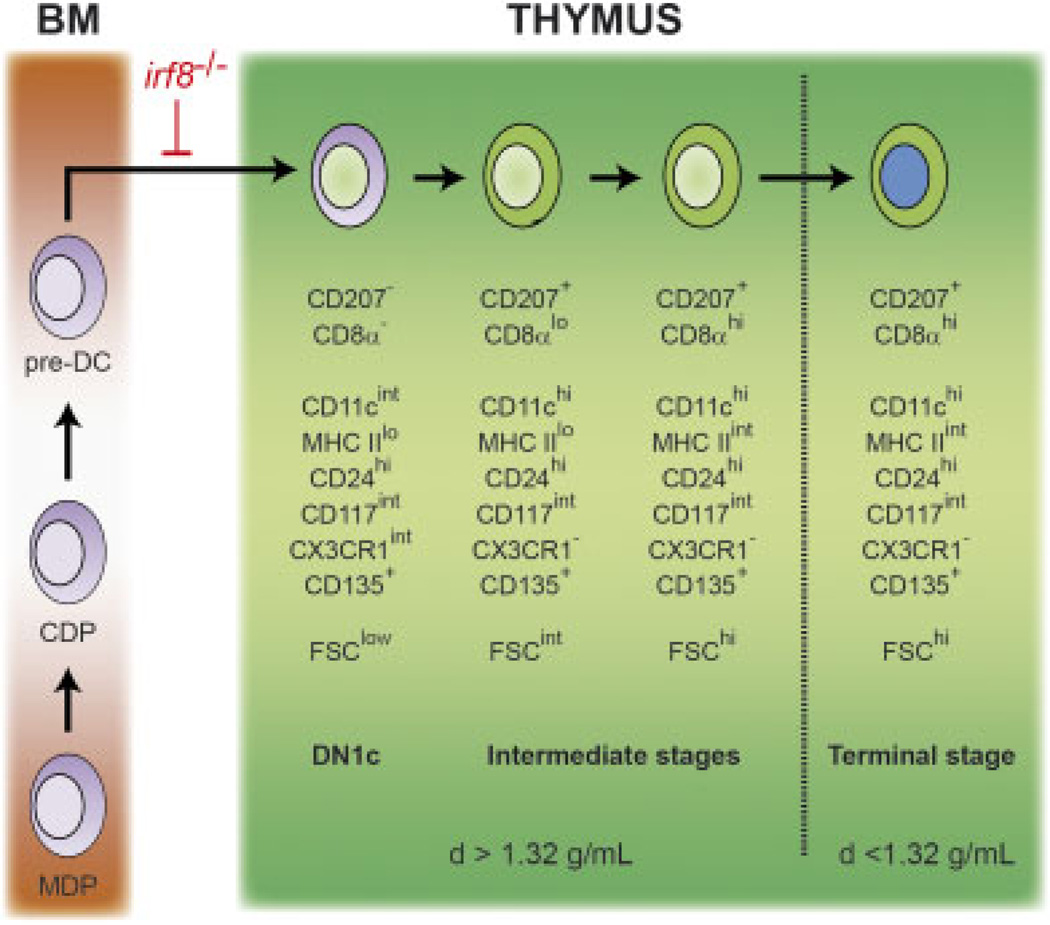
Comment in
-
A missing link in thymic dendritic cell development.Eur J Immunol. 2011 Aug;41(8):2145-7. doi: 10.1002/eji.201141850. Eur J Immunol. 2011. PMID: 21792873
References
-
- Petrie HT, Zuniga-Pflucker JC. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu. Rev. Immunol. 2007;25:649–679. - PubMed
-
- Klein L, Hinterberger M, Wirnsberger G, Kyewski B. Antigen presentation in the thymus for positive selection and central tolerance induction. Nat. Rev. Immunol. 2009;9:833–844. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
