

Reading TE leaves: new approaches to the identification of transposable element insertions
- PMID: 21632748
- PMCID: PMC3106314
- DOI: 10.1101/gr.110528.110
Reading TE leaves: new approaches to the identification of transposable element insertions
Abstract
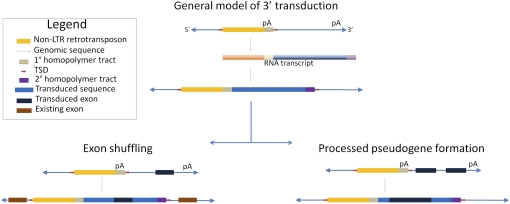
Transposable elements (TEs) are a tremendous source of genome instability and genetic variation. Of particular interest to investigators of human biology and human evolution are retrotransposon insertions that are recent and/or polymorphic in the human population. As a consequence, the ability to assay large numbers of polymorphic TEs in a given genome is valuable. Five recent manuscripts each propose methods to scan whole human genomes to identify, map, and, in some cases, genotype polymorphic retrotransposon insertions in multiple human genomes simultaneously. These technologies promise to revolutionize our ability to analyze human genomes for TE-based variation important to studies of human variability and human disease. Furthermore, the approaches hold promise for researchers interested in nonhuman genomic variability. Herein, we explore the methods reported in the manuscripts and discuss their applications to aspects of human biology and the biology of other organisms.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources